
Late Holocene foraminiferal record from mangrove reserve, Qi’ao Island, Pearl River Estuary and its implications for paleoenvironment
-
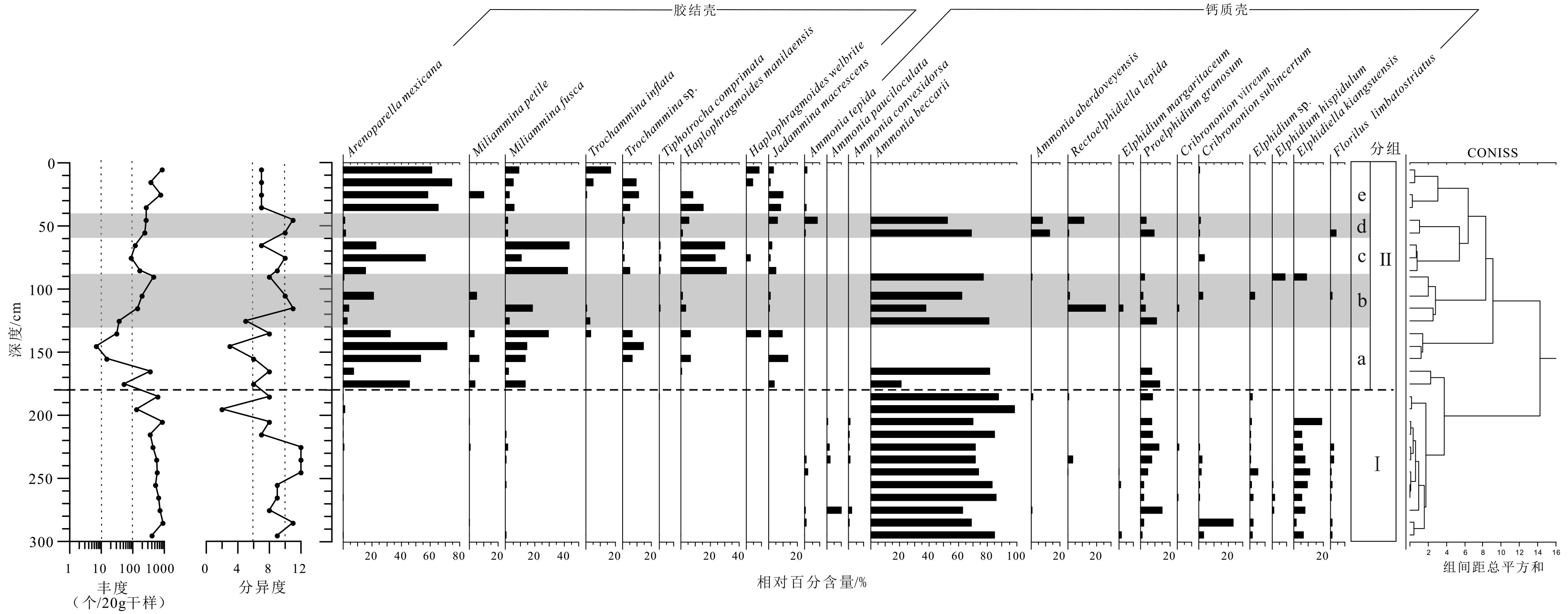
摘要: 潮间带有孔虫近年来被广泛地用于高精度古海平面等环境变化的研究中,但是在华南沿海此类研究少见。本文选取珠江口淇澳岛红树林潮间带上部长度为300 cm的QA钻孔进行了AMS14C测年,有孔虫、沉积物粒径、灰度分析,初步探究了研究区有孔虫作为高精度海平面指标的可能性及其所反映的古环境意义。研究结果显示钻孔包含两种有孔虫组合,一种是钙质有壳组合,常见属种为Ammonia becсarii(78.7%),Proelphidium granosum(6.6%)和Elphidiella kiangsuensis(5.6%),沉积底质为黏土质粉砂,对应沉积环境为河口潮下带;另一种是胶结壳组合,常见属种为Arenoparella mexicana(50.6%)、Miliammina fusca(17.5%)、Trochammia sp.(6.8%),沉积底质为砂质粉砂,含丰富植物碎屑,对应沉积环境为潮间带中上部红树林潮滩。结合AMS14C结果,QA钻孔记录了约4 200年以来的沉积,研究区初始为河口浅滩,由于三角洲进积作用浅滩淤积至海平面以上,约2 400年前研究区形成红树林中高潮滩。红树林沉积层中夹有两层粒度较细的钙质壳有孔虫沉积,推测可能由区域性次一级的海平面上升导致。灰度值反映了红树林沉积层中3个有机质含量较高的阶段,指示了温暖潮湿的气候,年代大约为3 000~2 700 cal. aBP, 1 200~1 100 cal. aBP, 600~500 cal. aBP,与海平面上升阶段较为接近。Abstract: Intertidal foraminifera has been widely used in paleoenvironmental reconstruction, and especially, in researches on high-resolution relative sea-level fluctuation. Unfortunately, such researches are lacking in the South China coast. This time, a long core up to 300 cm in length was retrieved from the upper intertidal zone of the mangrove on the Qi’ao Island of the Pearl River estuary. AMS14C dating, foraminifera, grain size distribution and grayscale analysis were employed to ascertain the possibility to apply the mangrove foraminifera as a high-resolution proxy for sea level fluctuation and paleoenvironmental evolution. Two foraminiferal assemblages were acquired from the core QA, a calcareous group dominated by Ammonia becсarii (78.7%), with common species Proelphidium granosum (6.6%) and Elphidiella kiangsuensis (5.6%), from a clayey slit sediment, indicating an estuarine subtidal environment; and an agglutinated assemblage dominated by Arenoparella mexicana (50.6%) with common species Miliammina fusca (17.5%), Trochammia sp. (6.8%), from a layer of sandy silt deposits with abundant plants debris and residues, indicating an upper or middle mangrove intertidal environment. AMS14C dates suggest that, the sediments formed approximately 4 200 cal. aBP, when the study area was a subtidal or mudflats environments. It turned to a middle-high mangrove at ~2 400 cal. aBP, resulted from the falling of relative sea-level caused by deltaic progradation. Two fine grained layers were intercalated in mangrove sediments, which contain calcareous foraminiferal tests corresponding to 1 022 ~ 729 cal. aBP and 508 ~ 358 cal. aBP respectively, probably related to a sub-regional sea-level rise. The grayscale suggests three phases of high humification, corresponding to ~3 000~2 700 cal. aBP, 1 200~1 100 cal. aBP, 600~500 cal. aBP respectively, indicating a warm and humid climate, which are close to the sea-level rise periods.
-
Keywords:
- mangrove /
- intertidal foraminifera /
- relative sea-level changes /
- paleoenviroment /
- the Qi’ao Island
-
潮滩是在潮汐作用显著、细颗粒沉积物供应丰富条件下形成的一种地貌类型,位于陆地、海洋、大气及人类活动相互作用的敏感地带。潮滩广泛分布在除基岩海岸和高纬度(70°~73°以上)海岸外的沿海地区,并且随着潮差、波浪、物质供应、植被类型等不同而表现出显著的空间差异[1-4]。目前全球潮滩面积约为127921 km2,主要分布在亚太地区,如印尼、中国、印度等国家[5]。我国的潮滩规模大且分布广泛,其中长江口-杭州湾及以北沿海地区主要为平原型潮滩,而长江口-杭州湾以南的浙、闽、粤、桂等沿海地区主要为港湾型潮滩[6]。从20世纪初开始,各国学者对潮滩沉积和地貌特征、潮滩沉积物输运和堆积的过程、盐沼植被对潮滩沉积过程的影响、极端事件对潮滩地貌演化影响等方面进行了系统研究和总结[7-16]。进入21世纪,随着现场观测技术和数值模拟的不断改进,我国科研人员开始关注潮滩沉积过程及其控制机制、植被-泥沙-地貌耦合的生物地貌过程、极端事件影响下的潮滩沉积物输运与地貌演化过程、潮滩沉积层序成因机制及潮滩演化趋势模拟等方面,取得了丰硕的成果[17-33]。
潮滩作为重要的海岸带生态系统,不仅在蓝色碳汇方面发挥着重要作用[34-35],而且在海岸防护方面起到至关重要的作用[36]。然而,随着海平面不断上升、地面沉降、流域及海岸带人类活动的不断加剧,潮滩面积逐渐减少[5],其中人类活动是主要驱动因素[37]。潮滩面积的减少不仅导致蓝色碳汇能力减弱[5, 35],而且引起诸多地区海岸遭受不同程度侵蚀[38]。高强度的人类活动显著影响了潮滩局部沉积物输运过程及地貌演化过程[20, 24],而流域输沙量的锐减则引起了潮滩冲淤格局的改变[39]。随着流域及河口地区人类活动强度的不断增大,河口系统状态正在发生转换[40],直接影响着河口潮滩未来演化趋势及其功能发挥。
本文选择我国东南沿海典型的山溪性中小型河流河口潮滩作为研究区域,通过沉积物采样、定点冲淤观测和沉积动力学观测等手段,初步分析了河口潮滩的季节性冲淤变化过程及其控制机制,以期为科学评估潮滩的海岸防护和潮滩生态系统保护与修复能力提供科学依据。
1. 研究区概况
闽江发源于福建与江西省交界的武夷山东麓及仙霞岭南麓,总体呈NW-SE向横贯福建中北部,经福州汇入东海南部,全长2872 km,其中干流全长近577 km,流域面积为60992 km2,多年平均径流量为605.5×108 m3,多年平均输沙量为750×104 t,并且表现出明显的洪、枯季变化,即洪季(3—8月)径流量约占全年的76%,输沙量约占全年的92%[41]。近年来随着流域水库建设的不断增加,平均输沙量已经下降为原来的三分之一[42]。
受地转偏向力及台湾海峡地形效应作用,闽江河口区潮汐作用强,实测最大潮差达到7.04 m,平均潮差4.46 m(梅花站),属正规半日潮,河口内潮流基本呈往复流,其流向与河槽线一致,河口外潮流具有一定的旋转流特征[41]。闽江口外海面开阔,夏季以西南风为主,冬季以东北风为主,波浪作用强,波浪的主要形式为风浪及涌浪同时存在的混合浪,多年平均波高为1.1 m,历史最大波高为6.5 m[41]。此外,闽江口海域是我国东南沿海地区遭受台风影响最为严重的区域之一,平均每年影响的台风次数为5.7个,最大台风增水值可达2.52 m[43]。
闽江河口地区的潮滩主要分布在川石岛南部、琅岐岛东部以及梅花水道南侧海岸(图1a),其中琅岐岛潮滩宽度为600~1100 m(图1b),滩面平缓。现场调查结果显示(图1c),琅岐岛潮滩由岸向海主要分布有互花米草、海三棱藨草、光滩等,其中互花米草盐沼滩面平均坡度约为0.36°,海三棱藨草盐沼滩面平均坡度约为0.14°,潮间带中下部的光滩平均坡度约为0.08°,河道边缘地区的滩面坡度约为1.84°。
![]() 图 1 研究区位置(a)、沉积物采样站位(b)、剖面形态及观测站位图(c)Figure 1. Sketch map of study area (a), sediment sampling sites (b), morphological character of profile and observation sites of tidal flat of Langqi Island (c)
图 1 研究区位置(a)、沉积物采样站位(b)、剖面形态及观测站位图(c)Figure 1. Sketch map of study area (a), sediment sampling sites (b), morphological character of profile and observation sites of tidal flat of Langqi Island (c)2. 研究方法
2.1 野外采样与观测
在琅岐岛东部潮滩选择一条剖面利用RTK对剖面进行了滩面高程测量,并在剖面上由岸向海设置了10个采样站位(图1b),分别于2022年8月和2023年2月采集了表层1 cm厚度的沉积物。为了掌握潮间带不同空间位置的冲淤变化特征,在互花米草盐沼边缘(L03站)、海三棱藨草盐沼中部(L05站)及光滩外缘(L09站)设置了3个站位,安装了冲淤观测架,在观测架上设置12个点,定期测量每个点距离滩面的高度,然后对各站位每次的测量数据进行平均,获得各站位的冲淤变化信息(图1c)。
为了解研究区水动力状况,在海三棱藨草内部靠近互花米草边缘设置一个沉积动力学观测站位(图1c,L04站),利用Nortek公司生产的声学多普勒海流计(ADV,VECTOR 6MHz)分别于2022年1月25日至2月1日、7月21—31日开展近底部潮流观测,仪器探头距离滩面30 cm,观测点位于滩面以上15 cm处;观测设置为脉冲模式进行采样,采样间隔为10 min,其中冬季采样频率为4 Hz,每次采集1024组数据,夏季采样频率为16 Hz,每次采集4096组数据;冬季利用北京海洲赛维科技有限公司生产的波潮仪(TWaves)进行淹没水深观测,以脉冲模式进行采样,以10 min为采样间隔,采样频率设置为8 Hz,每次采集时长为5 min,取平均值。
2.2 样品分析与数据处理
2.2.1 沉积物粒度分析
在实验室内将采集回来的常温保存样品充分混合,取约2 g重的沉积物加入浓度为5%的H2O2溶液以去除有机质,再加入浓度为1 mol/L的HCl溶液以去除碳酸盐,用蒸馏水清洗至溶液为中性,加入浓度为5‰的六偏磷酸钠溶液((NaPO3)6)浸泡24 h,并利用超声波震荡1 min,使样品充分混和、分散后,用英国马尔文公司生产的 Mastersizer 2000型激光粒度仪进行粒度分析,并用仪器自带软件导出中值粒径和各粒级组分数据,采用谢帕德分类体系对沉积物进行分类[44]。
2.2.2 悬沙浓度校对
利用在L04站采集的表层沉积物在室内配置不同悬沙浓度(SSC)的水体与ADV采集的声学信号进行相关分析,结果发现两者之间具有很好的相关性(图2)。因此,根据该回归方程可以将ADV观测获得的信噪比(Signal-Noise Ratio,SNR)转化成时间序列的SSC。
2.2.3 沉积动力学参数计算
利用仪器自带软件将观测数据导出,然后再利用该软件的数据转换功能,将原始数据转换成数据文件以便计算水平方向上的平均流速和波浪参数。
潮间带地区作为极浅水环境,整个水层都属于边界层范围内,平均水平流速在垂向上的分布遵循对数分布[45]:
$$ U\left(z\right)=\frac{{u}_{\mathrm{*}}}{\kappa}\mathrm{l}\mathrm{n}\left(\frac{z}{{z}_{0}}\right) $$ (1) $$\tau_{\rm b} = \rho_{w}u_{*}^{2} $$ (2) 其中,U(z)是平均水平流速度,u*是摩阻速度;κ是卡门常数(κ=0.4);z是距底床高度,z0是底部粗糙长度,τb是潮流引起的底部切应力,ρw是海水的密度。一般在边界层范围内,可以利用对数剖面方法计算潮流引起的底部切应力。
有效波高(Hs)采用如下公式计算[46]:
$$ H_{\rm s}=4\sqrt{\int S_ {\eta}(f){\rm d}f} $$ (3) 式中,Sη是水位的功率谱。将ADV测得的压力转换到水位时,需要考虑衰减,根据线性波理论,衰减系数可以表示为[47]:
$$ K f=\frac{\cosh [k(z+h)]}{\cosh k h} $$ (4) 其中,k是波数,h是平均水深,z是压力传感器的深度(负值)。波浪引起的底部切应力(τw)可以用如下公式计算[45]:
$$ \tau_{\rm w} = 0.5\rho f_{w}U_{w}^{2} $$ (5) 其中,fw为波浪摩擦因子,Uw为波浪轨迹速度,可以表达为波高(Hs)、波周期(T)和水深(h)的函数[45]:
$$ U_{\rm w} =\pi H_{\rm s} / [T{\rm{sinh}}(kh)] $$ (6) 其中k为波数,可以表达为:k = 2π/L,L为波长,可以用图解法计算得到[48]。波浪摩擦因子可以表达为:
$$ f_{w} = 1.39(A/z_{0})-0.52 $$ (7) 式中A为半轨迹距离,可以表达为:A=UwT/2π。
由流速和悬沙浓度数据计算近底层悬沙瞬时水平输运率f (t)及潮周期内悬沙净水平输运通量F (t)[45]:
$$ f(t) = U(z, t)\times C(z, t) $$ (8) $$ F(t) = \int_0^T {f(t)} {\rm d}t = \sum\limits_1^j {\Delta t{f_j}} (t) $$ (9) 其中U(z, t)和C(z, t) 分别表示距离海底高度z(m)处t时刻的瞬时水平流速(ua,m/s)和悬沙浓度 (SSC, mg/L),Δt表示实测时刻代表时间长度。
研究区表层沉积物冬季以粗颗粒沉积物为主,夏季以细颗粒沉积物为主,因此,在计算沉积物临界侵蚀切应力时分别选择适合非粘性沉积物和粘性沉积物的计算公式。非粘性沉积物临界侵蚀切应力时选择如下计算公式[45]:
$$ \theta = {\tau _{\text{b}}}/\left[ {{\text{g}}\rho \left( {s - 1} \right)d} \right] $$ (10) $$ {\theta _{{\text{cr}}}} = \frac{{0.30}}{{1 + 1.2{D_*}}} + 0.055\left( {1 - {{\rm e}^{ - 0.020{D_*}}}} \right) $$ (11) $$ {D_{\text{*}}} = {\left[ {{\rm g}\left( {s - 1} \right)/{\nu ^2}} \right]^{1/3}}d $$ (12) 其中,θ是Shields参数,θcr是沉积物临界起动Shields参数,s=ρs/ρw,ρs是沉积物密度(取值为2650 kg/m3),ρw是海水密度(取值为1025 kg/m3),d是沉积物粒径,g是重力加速度(取值为9.81 m/s2), D*是一个与沉积物粒径有关的无量纲参数,ν是水体的运动黏滞系数,取值为1.36×10−6 m2/s。对于黏性沉积物,其临界侵蚀切应力计算用如下公式[49]:
$$ {\tau _{\rm{cr}}} = 0.05 + \beta {\left\{ {\frac{1}{{{{\left[ {({\text π} /6)(1 + sW)} \right]}^{1/3}} - 1}}} \right\}^2} $$ (13) 其中,β是与颗粒粒径有关的系数,根据试验资料[49],在中值粒径小且表层沉积物含水量较大时,取β=0.3,W为含水量,定义为样品中水的质量与沉积物质量之比,根据采集的现场表层底质样品的分析获得。
3. 研究结果
3.1 沉积物粒度的季节分布特征
粒度分析结果显示(图3、图4),琅岐岛潮滩表层沉积物粒度组分以粉砂和砂为主,黏土含量相对较少,呈现出显著的时空差异。从季节方面看,冬季表层沉积物砂含量显著增大,黏土含量显著减小,沉积物中值粒径Φ值较小;夏季则相反,砂含量显著减小,黏土含量和粉砂明显增大,沉积物中值粒径Φ值较大(表1)。从空间上看,潮间带上部的互花米草盐沼内表层沉积物组分以粉砂为主,平均含量为60.6%,砂含量平均为21.0%,黏土含量平均为18.4%,中值粒径平均为5.60 Φ;在潮间带中部的海三棱藨草盐沼,沉积物组分以砂为主,平均含量为57.1%,粉砂含量平均为32.4%,黏土含量平均为10.5%,中值粒径平均为4.31 Φ;在潮间带下部的光滩地区,表层沉积物组分以粉砂为主,平均含量为58.7%,砂含量平均为23.0%,黏土含量平均为18.3%,中值粒径平均为5.56 Φ(图3、图4)。
![]() 图 3 琅岐岛潮滩夏季(左)和冬季(右)表层沉积物粒度组成Figure 3. Surficial sediment composition along the profile of tidal flat in summer (left) and winter (right)
图 3 琅岐岛潮滩夏季(左)和冬季(右)表层沉积物粒度组成Figure 3. Surficial sediment composition along the profile of tidal flat in summer (left) and winter (right)![]() 图 4 琅岐岛潮滩不同季节表层沉积物中值粒径Figure 4. Seasonal variations of median grain-size of surficial sediment along the profile of tidal flat表 1 琅岐岛潮间带不同季节表层沉积物粒度组分及中值粒径Table 1. Seasonal variations of composition and median grain-size of surficial sediment
图 4 琅岐岛潮滩不同季节表层沉积物中值粒径Figure 4. Seasonal variations of median grain-size of surficial sediment along the profile of tidal flat表 1 琅岐岛潮间带不同季节表层沉积物粒度组分及中值粒径Table 1. Seasonal variations of composition and median grain-size of surficial sediment采样时间 砂含量/% 粉砂含量/% 黏土含量/% 中值粒径/Φ 变化范围 平均值 变化范围 平均值 变化范围 平均值 变化范围 平均值 2022年8月 0.2~39.9 10.3 43.6~70.3 63.6 16.4~32.9 26.1 5.03~7.23 6.52 2023年2月 14.6~95.7 60.8 4.0~73.3 32.8 0.2~14.4 6.4 2.57~5.24 3.79 根据Shepard三角分类图,琅岐岛潮间带表层沉积物类型以黏土质粉砂(YT)为主,并且呈现出显著的季节变化,其中夏季表层沉积物黏土质粉砂占80%,仅在潮间带中部出现砂质粉砂(ST)和砂-粉砂-黏土(STY);冬季以砂(S)和砂质粉砂为主,其次为粉砂质砂(TS)(图5)。
![]() 图 5 琅岐岛潮滩不同季节表层沉积物类型Figure 5. Seasonal variations of surficial sediment type along the profile of tidal flat based on Shepard classification
图 5 琅岐岛潮滩不同季节表层沉积物类型Figure 5. Seasonal variations of surficial sediment type along the profile of tidal flat based on Shepard classification3.2 潮滩侵蚀-淤积的时空变化特征
潮间带不同空间位置的冲淤观测结果显示,琅岐岛潮间带滩面冲淤表现出显著的时空变化特征。互花米草盐沼边缘滩面表现出总体持续淤积的变化特征,仅在冬末至初春季节出现轻微侵蚀(0.55 cm),观测期间年淤积厚度为1.22 cm(图6a)。海三棱藨草中部滩面在初夏至初秋季节表现为持续淤积,淤积厚度为14.13 cm;初秋至冬末季节表现为持续侵蚀,侵蚀深度为20.13 cm;冬末至春末期间呈现出缓慢淤积,淤积厚度为2.54 cm;整个初夏至翌年春末表现为净侵蚀,侵蚀深度为3.42 cm(图6b)。光滩外缘滩面在初夏至秋末季节表现为持续侵蚀,侵蚀深度为5.90 cm;秋末至翌年春末季节表现为持续淤积,侵蚀深度为5.38 cm,整个初夏至翌年春末表现为净侵蚀,侵蚀深度为0.52 cm(图6c)。
![]() 图 6 潮间带不同站位冲淤变化过程正值为淤积,负值为冲刷。Figure 6. Variations of erosion and accretion along the section in different stationsPositive value means accretion, negative value means erosion.
图 6 潮间带不同站位冲淤变化过程正值为淤积,负值为冲刷。Figure 6. Variations of erosion and accretion along the section in different stationsPositive value means accretion, negative value means erosion.3.3 潮间带水动力特征
琅岐岛潮间带中上部L04站位淹没水深观测结果显示(图7a、8a),冬季和夏季观测期间大潮最大淹没水深分别为2.20和1.86 m,小潮最大淹没水深分别为1.37和0.90 m。潮流观测结果显示(图7b、8b),冬季小潮观测期间近底部潮流流速为0.008 ~0.127 m/s,大潮期间为0.004 ~0.187 m/s;夏季小潮观测期间近底部潮流流速为0.001 ~0.046 m/s,大潮期间为0.001 ~0.060 m/s;无论冬季还是夏季,潮周期内潮流流速最大值均出现在落潮后期。波浪计算结果显示(图7c、8c),冬季和夏季观测期间有效波高最大值分别为0.63和0.20 m,平均值分别为0.26 和0.07 m,冬季波浪明显较夏季强。底部切应力计算结果显示(图7d、8d),冬季和夏季观测期间潮流引起的底部切应力最大值分别为0.551和0.057 N/m2,平均值分别为0.067和0.003 N/m2;波浪引起的底部切应力最大值分别为1.885和0.293 N/m2,平均值分别为0.652和0.097 N/m2,冬季潮流和波浪引起的底部切应力明显大于夏季。
![]() 图 7 潮滩中上部光滩冬季近底部水动力时间序列变化过程a:淹没水深,b:近底部流速,c:有效波高,d:底部切应力,e:近底部悬沙浓度。Figure 7. Time-series of near-bottom hydrodynamic processes in winter on the mid-upper tidal flata: Inundation height, b: near-bottom mean current speed, c: significant wave height, d: bottom shear stress induced by tidal current and wave, e: near-bottom suspended sediment concentration.
图 7 潮滩中上部光滩冬季近底部水动力时间序列变化过程a:淹没水深,b:近底部流速,c:有效波高,d:底部切应力,e:近底部悬沙浓度。Figure 7. Time-series of near-bottom hydrodynamic processes in winter on the mid-upper tidal flata: Inundation height, b: near-bottom mean current speed, c: significant wave height, d: bottom shear stress induced by tidal current and wave, e: near-bottom suspended sediment concentration.3.4 潮间带近底部悬沙浓度变化
悬沙浓度计算结果显示,琅岐岛潮间带中上部L04站位近底部悬沙浓度呈现出显著的时间变化特征(图7e、8e)。在季节尺度上,冬季观测期间近底部悬沙浓度为17.4 ~844.5 mg/L,平均值为306.8 mg/L,而夏季观测期间近底部悬沙浓度为3.4 ~607.8 mg/L,平均值为138.5 mg/L,冬季悬沙浓度显著高于夏季。在大-小潮周期尺度上,小潮期间近底部悬沙浓度相对较低,大潮期间相对较高,但潮周期平均值相差不大,如冬季大潮和小潮观测期间近底部悬沙浓度平均值分别为296.5和268.6 mg/L,而夏季大潮和小潮观测期间近底部悬沙浓度平均值分别为158.1和123.7 mg/L。在潮周期尺度上,涨潮初期和落潮后期相对较高,高平潮期间悬沙浓度相对较低。
3.5 潮间带近底部悬沙输运
近底部悬沙东向水平输运率计算结果呈现出显著的时间变化特征(图9),冬季观测期间近底部悬沙在东向上水平输运率为−0.118 ~0.051 kg·m−2·s−1,平均值为0.011 kg·m−2·s−1;夏季观测期间近底部悬沙水平输运率为−0.006 ~0.009 kg·m−2·s−1,平均值为0.001 kg·m−2·s−1。
![]() 图 9 近底部瞬时悬沙水平输运率及潮周期内悬沙水平输运净通量a:冬季,b:夏季。正值为向海输运,负值为向岸输运;T1—T19为观测期间的潮周期数。Figure 9. Seaward transport rate and net flux of near-bottom instantaneous suspended sediment in a tidal cycle in winter (a) and summer (b)Positive values indicate seaward transport, negative ones are landward transport; T1—T19: the number of tidal cycle during the observation.
图 9 近底部瞬时悬沙水平输运率及潮周期内悬沙水平输运净通量a:冬季,b:夏季。正值为向海输运,负值为向岸输运;T1—T19为观测期间的潮周期数。Figure 9. Seaward transport rate and net flux of near-bottom instantaneous suspended sediment in a tidal cycle in winter (a) and summer (b)Positive values indicate seaward transport, negative ones are landward transport; T1—T19: the number of tidal cycle during the observation.每个潮周期内的东-西方向上的净悬沙输运量计算结果显示(图9),琅岐岛潮间带中部潮周期内近底部悬沙水平净输运通量呈现出明显的时间变化特征。在大-小潮时间尺度上,小潮期间潮周期内悬沙净水平输运通量显著小于大潮,而净输运方向则呈现出季节性差异:小潮期间冬季呈现出向岸净输运和向海净输运交替出现,而夏季则整体呈现向海净输运的变化特征;中潮期间冬季总体表现为向海净输运,而夏季则整体呈现向陆净输运的变化特征;大潮期间冬季总体呈现向岸净输运,而夏季则呈现向岸净输运和向海净输运交替出现的变化特征。在季节尺度上,冬季潮周期内悬沙净水平输运通量为−124.30~243.24 kg/m,观测期间净输运通量为289.49 kg/m;夏季周期内悬沙净水平输运通量仅为−38.07 ~4.13 kg/m,观测期间净输运通量仅为−58.73 kg/m,即冬季潮周期内净水平输运通量显著大于夏季,并且冬季悬沙净向海输运,而夏季净向陆输运。
4. 讨论
河口潮滩是由地质过程以及水动力过程、沉积物供应和生态过程的相互作用形成的,其地貌形态及沉积物组成受动力、物源及生物活动控制,具有高度的时空变化特征[4, 50]。大量的河流入海泥沙进入河口后,在水动力的作用下发育了广泛的潮滩[6, 51],但由于我国河流泥沙入海主要集中在夏季,冬季入海泥沙通量很小[41],因此,潮滩表层沉积物也表现出显著的季节差异[52-54]。闽江河流泥沙入海通量也主要集中在夏季[55],本文断面表层沉积物分析结果(图3、图4)和冲淤观测结果(图6)也显示,夏季期间河口潮滩捕获了大量的河流入海沉积物,潮滩为淤积状态,潮滩表层沉积物以粉砂和黏土为主,平均粒径较小;冬季期间河流入海泥沙很少,河口潮滩很少能捕获到来自河流的泥沙,潮滩表层沉积物在水动力的作用下不断发生改造,最终细颗粒沉积物被搬运带走,潮滩发生侵蚀,表层沉积物以砂和粉砂为主。关于沉积物供应变化对闽江口潮滩沉积物组成的影响综合分析详见李海琪等[56],本文不再赘述,但需要指出的是,随着闽江入海泥沙通量的显著减少[55],河流输沙季节变化对琅岐岛潮滩季节性冲淤变化的影响将会减小。本文将主要从水动力方面阐述闽江河口潮滩冲淤的季节性变化机制。
影响潮滩沉积和地貌演化的动力过程主要包括潮汐、波浪、跨岸/沿岸流、风致环流、潮不对称等[50, 57],尤其是在极浅水环境下,潮流、波浪是影响潮滩沉积物分布的主要动力因素[27-28]。沉积物是否发生侵蚀和淤积,主要是看水动力引起的底部切应力是否会超过临界切应力,当水动力引起的底部切应力大于沉积物的临界侵蚀切应力,则沉积物发生再悬浮,滩面呈现侵蚀状态;当水动力引起的底部切应力小于泥沙临界沉降切应力,则泥沙发生沉降,滩面呈现淤积状态。根据本文观测期间采集的表层沉积物粒度分析结果,冬季表层沉积物砂含量达到了93.9%,中值粒径为2.85 Φ,属于典型的非粘性沉积物,而夏季表层沉积物砂、粉砂和黏土含量分别为24.4%、54.2%和21.4%,中值粒径为6.17 Φ,属于典型的粘性沉积物。因此,根据相应的计算公式可知,观测站位冬季和夏季表层沉积物临界侵蚀切应力分别为0.158和0.107 N/m2。对比结果发现(图7d、8d),琅岐岛潮滩中上部地区近底层流速相对较小,由此产生的底部切应力也相对较小,对沉积物的活动性影响不明显,但冬季观测期间波浪引起的底部切应力显著大于表层沉积物的临界侵蚀切应力,表明冬季观测期间观测站位附近滩面发生强烈的侵蚀;夏季观测期间波浪引起的底部切应力的变化大部分时刻均小于表层沉积物临界侵蚀切应力,表明夏季观测期间观测站位附近滩面表层沉积物不易发生侵蚀。在长江口及江苏沿海潮间带观测结果也显示,在这种极浅水环境下波浪作用显著影响潮滩冲淤过程[28, 58]。研究表明[59],悬沙临界沉降切应力(τcrd)一般为0.06~0.1 N/m2,根据Whitehouse等的推荐,本文取τcrd=0.08 N/m2(其值适用于细颗粒泥沙),则对比结果显示(图8d),夏季观测期间大部分时刻由潮流和波浪引起的底部切应力小于临界沉降切应力,表明观测期间观测站位附近水体悬沙易发生沉降,导致滩面出现淤积。近底部悬沙净输运通量计算结果显示,冬季观测期间泥沙净向海输运,而夏季则净向岸输运。该结果表明,波浪是引起潮间带表层沉积物发生侵蚀的主要动力,而潮流则是搬运泥沙的主要动力。涨落潮不对称现象是河口海岸地区重要的水动力特征,是影响区域泥沙不对称输运的重要因素[60],在河口海岸地区泥沙输运和地貌演变过程中扮演着十分重要的角色[57, 61]。由图8和图9可知,冬季涨潮期间悬沙浓度总体大于落潮,但由于波浪引起的底部切应力显著大于悬沙临界沉降切应力,导致悬沙很少发生沉降,涨潮流带来的悬沙除部分被潮间带上部的互花米草盐沼植被捕获外,其他悬沙又在落潮流的作用下向海输运,而研究区无论冬季还是夏季,潮流均以落潮流占优,因此,落潮期间的底层悬沙输运通量总体大于涨潮,潮周期内表现为净向海输运;夏季,虽然是落潮流占优,但涨潮期间的底层悬沙浓度总体大于落潮,仅在大潮期间波浪作用较强时期出现落潮悬沙浓度大于涨潮的情况,因此涨潮期间的悬沙输运通量总体大于落潮,并且在多数时刻底部切应力小于临界沉降切应力,悬沙沿途发生沉降。该结论得到了本文现场冲淤观测结果的支持(图6),同时也与其他河口地区潮滩冲淤过程一致[52, 62]。
![]() 图 8 潮滩中上部光滩夏季近底部水动力时间序列变化过程a:淹没水深,b:近底部流速,c:有效波高,d:底部切应力,e:近底部悬沙浓度。Figure 8. Time-series of near-bottom hydrodynamic processes in summer on the mid-upper tidal flata: Iinundation height, b: near-bottom mean current speed, c: significant wave height, d: bottom shear stress induced by tidal current and wave, e: near-bottom suspended sediment concentration.
图 8 潮滩中上部光滩夏季近底部水动力时间序列变化过程a:淹没水深,b:近底部流速,c:有效波高,d:底部切应力,e:近底部悬沙浓度。Figure 8. Time-series of near-bottom hydrodynamic processes in summer on the mid-upper tidal flata: Iinundation height, b: near-bottom mean current speed, c: significant wave height, d: bottom shear stress induced by tidal current and wave, e: near-bottom suspended sediment concentration.本文观测结果显示,琅岐岛潮间带中部地区,冬季观测期间的潮流和波浪作用(图7b、c)均明显较夏季强(图8b、c),这主要一方面是由于研究区冬季主要受东北季风控制,而夏季主要受西南季风控制,冬季期间风速大于夏季,由此引起研究区冬季期间潮流和波浪作用较夏季强[43, 63];另一方面,则与潮间带地貌的季节性变化有关。现场调查发现,琅岐岛潮间带中部夏季被海三棱藨草覆盖,而冬季则变为光滩。现场观测和数值模拟结果均表明,盐沼植被具有显著的弱流、消波作用[64-66]。琅岐岛潮滩夏季由于生长了大面积的海三棱藨草,起到了一定的缓流作用,因此,夏季观测期间最大潮流流速和平均潮流流速均小于没有盐沼植被覆盖的冬季,波浪作用也是夏季显著小于冬季,表明琅岐岛潮间带海三棱藨草的生长具有一定弱流和显著的消波作用。根据不同站位冲淤变化对比分析(图6),在观测期间互花米草盐沼边缘滩面冲淤变化幅度很小,并且与其他两个站位的冲淤变化趋势没有明显的一致性,说明该地区滩面冲淤主要受水动力和互花米草的影响。位于海三棱藨草覆盖区域和光滩区域滩面冲淤变化总体表现为相反的变化趋势,这可能与海三棱藨草生长和消亡会引起潮间带中部地区冲淤变化有关,即在有海三棱藨草覆盖时,在动力-植被-泥沙的耦合作用下,涨潮带来的泥沙被植被捕获并沉积下来[30],退潮水体悬沙浓度降低,导致在光滩地区退潮水流携沙能力有所增强,在一定程度上可能会引起潮间带下部光滩的侵蚀;当没有植被覆盖时,潮间带中部遭受显著侵蚀,侵蚀下来的泥沙在被落潮流携带至河口的过程中沿途发生沉降,引起潮间带下部滩面淤积。上述分析表明,盐沼植被在潮滩冲淤变化过程和潮滩发育演变中发挥着重要作用。
综上所述,琅岐岛潮间带中上部潮滩冲淤变化的动力学机制为:冬季,由于潮间带中部的海三棱藨草消失,潮间带被水淹没期间的波浪作用强,由此引起的底部切应力显著大于表层沉积物的临界侵蚀切应力,表层沉积物发生再悬浮,再悬浮起来的泥沙在潮流的作用下向海输运,导致滩面发生侵蚀;夏季潮间带中部被海三棱藨草覆盖,潮流和波浪均减弱,由此引起的底部切应力在大部分时刻小于表层沉积物的临界侵蚀切应力和临界沉降切应力,水体中的悬沙易发生沉降,导致滩面发生淤积。
5. 结论
(1)琅岐岛潮滩表层沉积物粒度组分以粉砂和砂为主,黏土含量相对较少,呈现出显著的时空差异,夏季主要以粉砂和黏土组成的细颗粒沉积物为主,冬季则主要以砂和粉砂组成的粗颗粒沉积物为主。
(2)琅岐岛潮间带上部的互花米草盐沼总体以持续淤积为主,潮间带中部和下部表现出周期性的冲淤特征,总体以夏季淤积、冬季侵蚀为总特征,并且潮间带中部的冲淤变化幅度较潮间带下部大。
(3)沉积动力观测与计算结果表明,琅岐岛潮间带中上部近底部潮流流速相对较小,但波浪作用强,近底部悬沙浓度相对较高,其中冬季潮流流速、波浪作用及悬沙浓度均大于夏季,冬季潮周期内近底部悬沙以净向海输运为主,而夏季则以净向岸输运为主。
(4)琅岐岛潮滩冲淤变化主要受动力过程控制,冬季因潮间带海三棱藨草消失,波浪作用强,导致表层沉积物易发生再悬浮,再悬浮的泥沙在较强的潮流作用下净向海输运;夏季因潮间带有海三棱藨草生长,波浪作用较弱,水体中的悬沙易发生沉降,较弱的潮流将泥沙净向岸输运,导致潮间带发生淤积。
致谢:黄书仁、黄思添、余永泽、陈海煌、刘三善参与了野外采样工作,黄书仁参与了实验室样品分析,谨致谢忱!
-
![]()
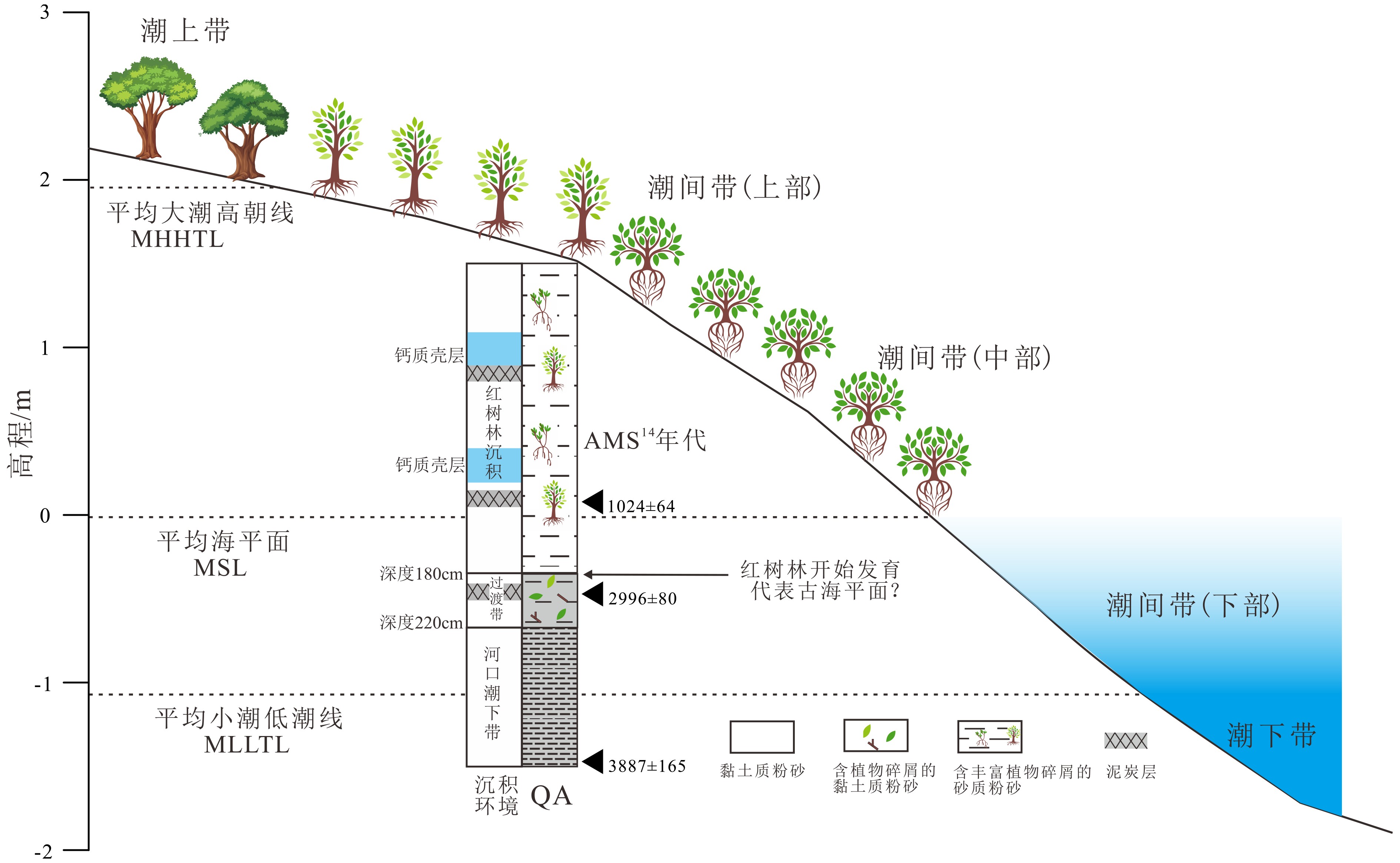
图 1 珠江口伶仃洋(a)以及淇澳岛钻孔位置(卫星图)(b)示意图
Figure 1. Map of Lingding Bay of Pearl River estuary (a) and location of core QA from Qi’ao Island in satellite image (b)
![]()
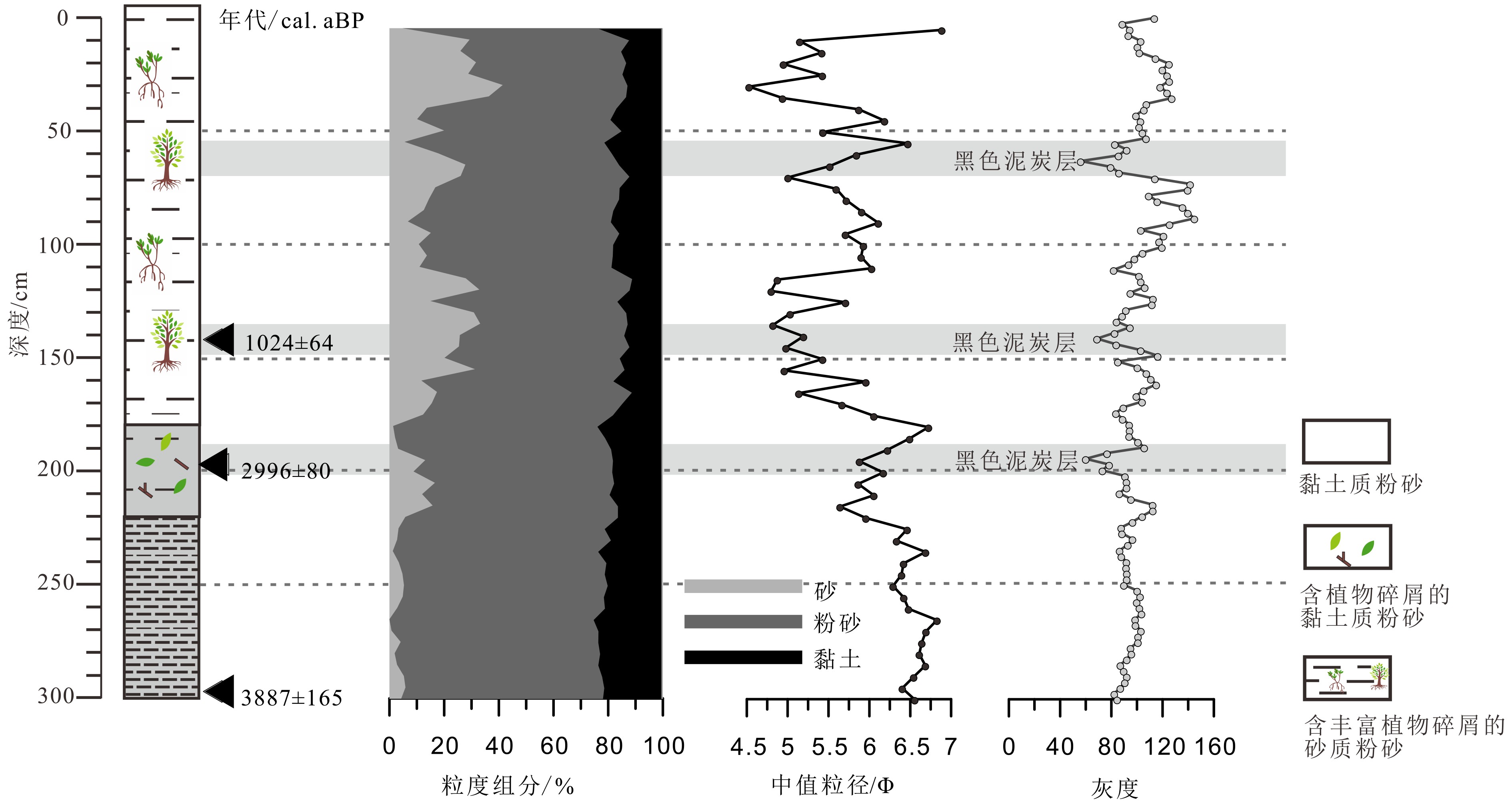
图 3 钻孔QA岩性、沉积物组分、灰度值、中值粒径随深度的变化曲线(灰色阴影表示3个深色泥炭层的位置)
Figure 3. Vertical variation of lithology, sediment fraction, gray scale, median grain size with depth from core QA ( The three gray shadow layers indicate peat horizons)
![]()
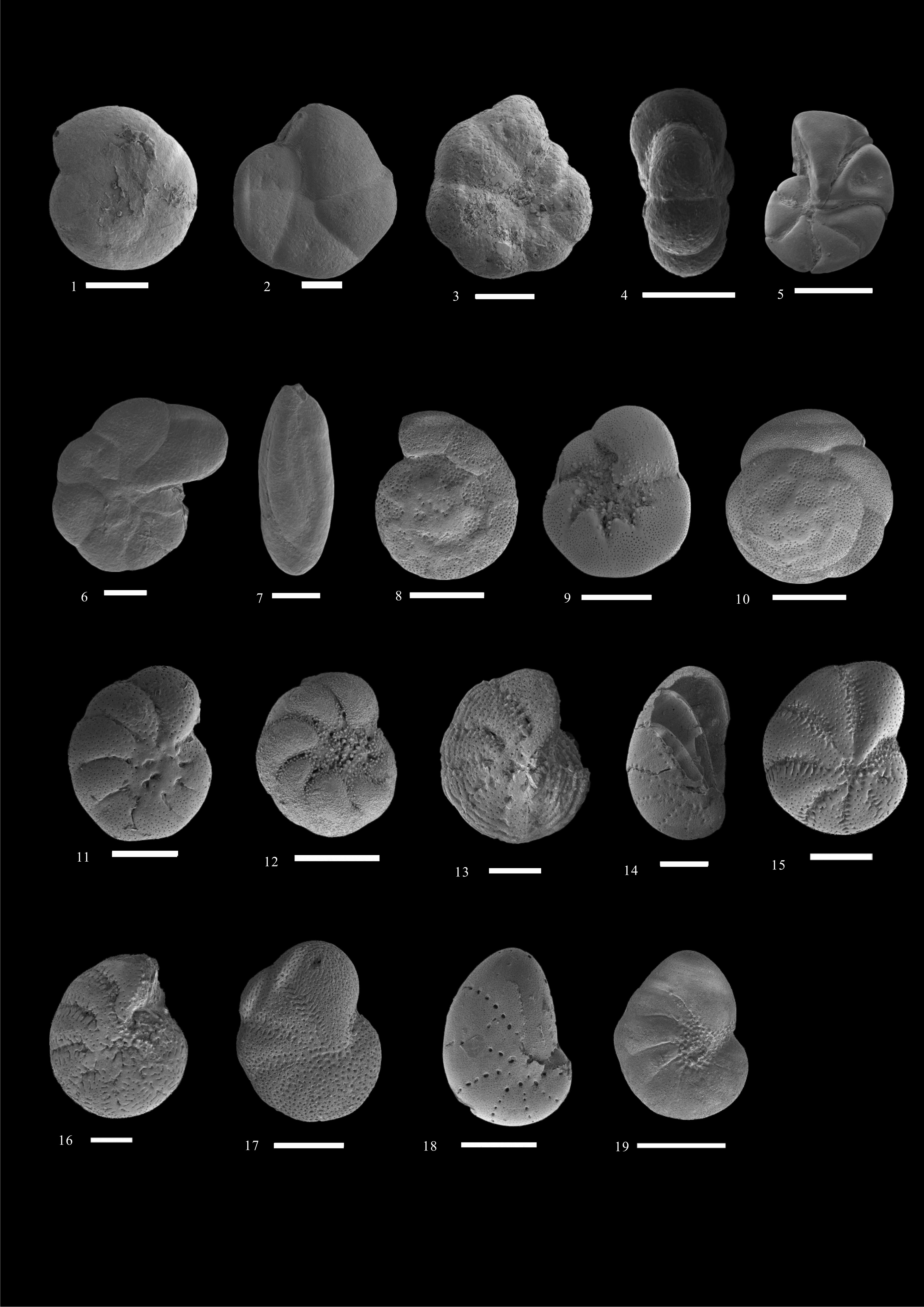
图 5 有孔虫扫描电镜照片 (比例尺=100 μm)
Figure 5. Scanning Electron Microscope pictures of foraminifera (scalebar=100 μm)
1,2. Arenoparrella mexicana (Kornfeld, 1931); 3,4. Haplophragmoides manilaensis (Andersen, 1952); 5. Trochammina sp. ;6. Jadammina macrescens (Brady, 1870); 7. Miliammina fusca (Brady, 1870); 8,9. Ammonia beccarii (Linné, 1758); 10. Ammonia aberdoveyensis (Haynes, 1973); 11. Cribrononion subincertum (Asano, 1951); 12. Proelphidium granosum (d’Orbigny, 1846); 13. Elphidium hispidulum (Cushman, 1936); 14. Rectoelphidiella lepida (He, Hu & Wang, 1965); 15. Elphidium sp. (Williamson, 1858) ;16. Elphidium margaritaceum (Cushman, 1930); 17. Elphidiella kiangsuensis (He, Hu & Wang, 1965); 18. Cribrononion vitreum (Wang, 1980); 19. Florilus limbatostriatus (Cushman, 1931)
![]()
![]()
图 7 淇澳岛红树林潮滩环境对应潮汐高程及古环境演变示意图(据QA孔)
Figure 7. Sketch map of tidal datum and mangrove tidal flat from Qi’ao Island, and its paleoenvironmental evolutions revealed by core QA
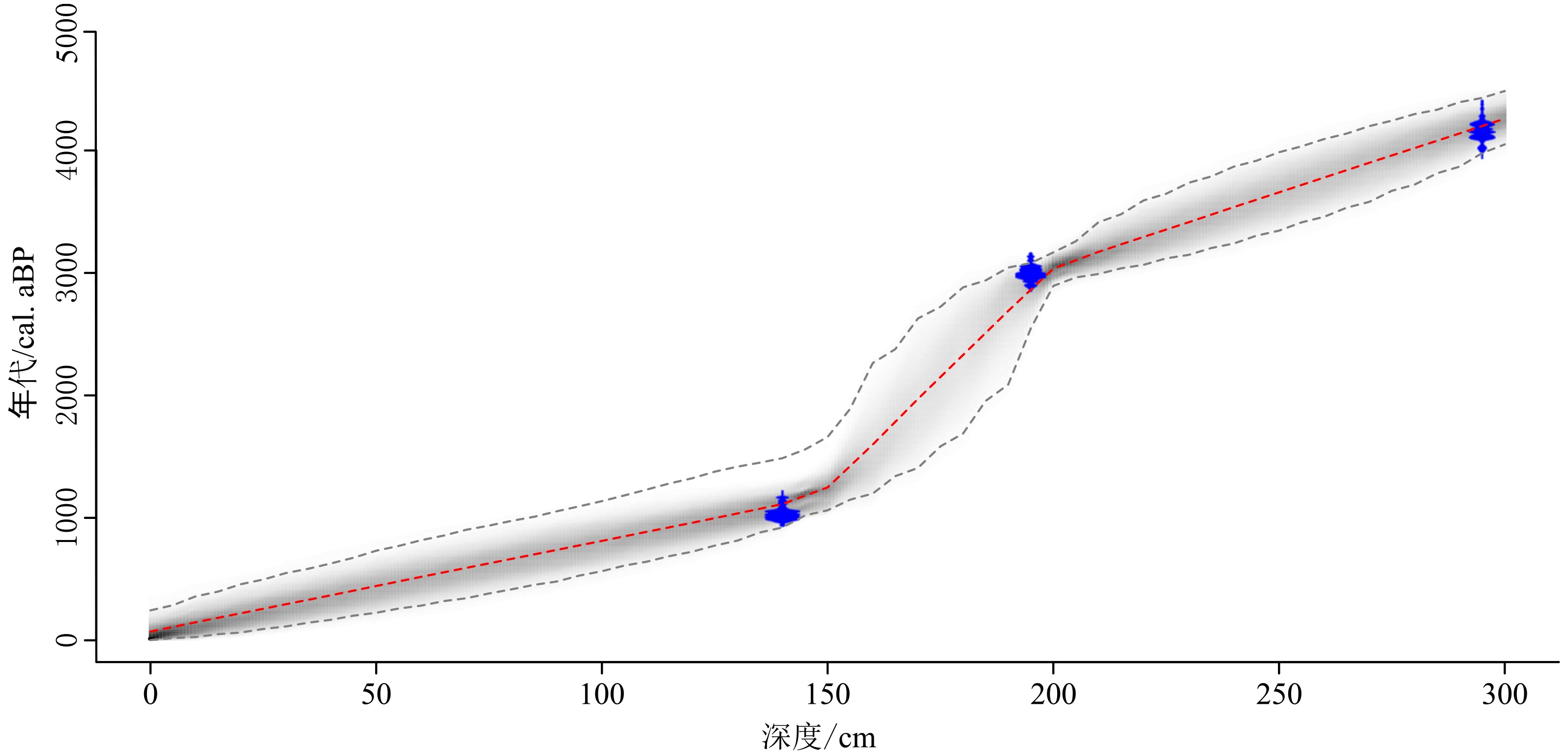
表 1 钻孔QA沉积物AMS14C测年结果
Table 1 AMS14C radiocarbon dates from sediment of core QA
样品编号 实验室编号 深度 /cm 测年材料 δ13C /‰ 14C年龄/a 校正年龄2σ /cal. aBP QA-C1 Poz#2-78133 140 贝壳 –10.2 1 124±28 1 024±64 QA-C3 Poz-78599 195 有机质泥 –26.7 2 868±30 2 996±80 QA-C2 Poz-78598 295 有机质泥 –23.6 3 779±32 3 887±165  下载: 导出CSV
下载: 导出CSV
-
[1] Frey R W, Basan P B. Coastal salt marshes[M]//Davis R A. Coastal Sedimentary Environments. New York, NY: Springer, 1985: 225-301.
[2] Macdonald K B. Quantitative studies of salt marsh mollusc faunas from the north American Pacific coast [J]. Ecological Monographs, 1969, 39(1): 33-60. doi: 10.2307/1948564
[3] 尤坤元, 朱大奎, 施晓冬. 潮汐盐沼环境特点及高分辨率海面变化有孔虫标尺[J]. 地理科学, 1999, 19(3):277-281. [YOU Kunyuan, ZHU Dakun, SHI Xiaodong. Environmental characteristics and high resolution indicators of foraminifer for change of sea levels in the tidal saltmarsh [J]. Scientia Geographica Sinica, 1999, 19(3): 277-281. doi: 10.3969/j.issn.1000-0690.1999.03.016 [4] Scott D S, Medioli F S. Vertical zonations of marsh foraminifera as accurate indicators of former sea-levels [J]. Nature, 1978, 272(5653): 528-531. doi: 10.1038/272528a0
[5] Horton B P, Edwards R. Quantifying Holocene sea level change using intertidal foraminifera: lessons from the british isles[C]. Philadelphia Annual Meeting, 2006.
[6] Kemp A C, Horton B P, Culver S J. Distribution of modern salt-marsh foraminifera in the Albemarle-Pamlico estuarine system of North Carolina, USA: Implications for sea-level research [J]. Marine Micropaleontology, 2009, 72(3-4): 222-238. doi: 10.1016/j.marmicro.2009.06.002
[7] Liu S M. Comparison of mangrove and salt marsh foraminifera as sea-level indicators [J]. Quaternary International, 2012, 279-280: 285.
[8] Woodroffe S, Horton B P, Larcombe P, et al. Intertidal mangrove foraminifera from the central Great Barrier Reef shelf, Australia: implications for sea-level reconstruction [J]. Journal of Foraminiferal Research, 2005, 35(3): 259-270. doi: 10.2113/35.3.259
[9] Horton B P, Larcombe P, Woodroffe S A, et al. Contemporary foraminiferal distributions of a mangrove environment, Great Barrier Reef coastline, Australia: implications for sea-level reconstructions [J]. Marine Geology, 2003, 198(3-4): 225-243. doi: 10.1016/S0025-3227(03)00117-8
[10] Debenay J P, Guiral D. Mangrove swamp foraminifera, indicators of sea level or paleoclimate? [J]. Revue De Paleobiologie, 2006, 25(2): 567-574.
[11] 李建芬, 商志文, 姜兴钰, 等. 渤海湾沿岸贝壳堤对潮滩有孔虫海面变化指示意义的影响[J]. 地质通报, 2016, 35(10):1578-1583. [LI Jianfen, SHANG Zhiwen, JIANG Xingyu, et al. Sea-level indicated by foraminifera assemblages liv-ing in the open muddy flats with or without influence of the chenier ridges in Bohai Bay coastal area [J]. Geological Bulletin of China, 2016, 35(10): 1578-1583. doi: 10.3969/j.issn.1671-2552.2016.10.003 [12] 王绍鸿, 喻鸣同, 唐丽玉, 等. 福建深沪湾潮间带全新世有孔虫及其环境意义[J]. 台湾海峡, 2002, 21(1):6-11. [WANG Shaohong, YU Mingtong, Tang Liyu, et al. Holocene foraminifera and it's environmental significance in Shenhu Bay, Fujian [J]. Journal of Oceanography in Taiwan Strait, 2002, 21(1): 6-11. [13] 尤坤元, 陈才俊, 吴小根, 等. 我国苏北建川潮汐盐沼有孔虫垂直分带的发现和特征[J]. 海洋学报, 2002, 24(1):59-64. [YOU Kunyuan, CHEN Caijun, WU Xiaogen, et al. Discovery and features of vertical zonations of tidal salt-marsh foraminifera in Jianchuan, North Jiangsu Province, China [J]. Acta Oceanologica Sinica, 2002, 24(1): 59-64. [14] 吴乃琴, 汪品先. 我国海岸带胶结壳有孔虫分布的控制因素[J]. 科学通报, 1989(12):924-927. [WU Naiqin, WANG Pingxian. The distribution and constraining factors of agglutinated foraminifera of Chinese coast [J]. Chinese Science Bulletin, 1989(12): 924-927. doi: 10.3321/j.issn:0023-074X.1989.12.001 [15] 李建芬, 商志文, 王宏, 等. 渤海湾西部现代有孔虫群垂直分带的特征及其对全新世海面、地质环境变化的指示[J]. 地质通报, 2010, 29(5):650-659. [LI Jianfen, SHANG Zhiwen, WANG Hong, et al. Modern foraminifera assemblages: vertical zonation and its indication for Holocene sea level and geoenvironmental reconstruction in Bohai Bay, China [J]. Geological Bulletin of China, 2010, 29(5): 650-659. doi: 10.3969/j.issn.1671-2552.2010.05.003 [16] 龙海燕. 胶州湾盐沼有孔虫分布特征及其环境意义研究[D]. 中国海洋大学博士学位论文, 2009. LONG Haiyan. Distribution of salt marsh foraminifera in Jiaozhou Bay and its environmental implication[D]. Doctoral Dissertation of Ocean University of China, 2009.
[17] 张留恩, 廖宝文. 珠海市淇澳岛红树林湿地的研究进展与展望[J]. 生态科学, 2011, 30(1):81-87. [ZHANG Liuen, LIAO Baowen. Research progress of mangrove wetlands on Qi’ao island, Zhuhai [J]. Ecological Science, 2011, 30(1): 81-87. doi: 10.3969/j.issn.1008-8873.2011.01.015 [18] 叶翔, 李靖, 王爱军. 珠江口淇澳岛滨海湿地沉积环境演化及其对人类活动的响应[J]. 海洋学报, 2018, 40(7):79-89. [YE Xiang, LI Jing, WANG Aijun. Sedimentary environment and its response to anthropogenic impacts in the coastal wetland of the Qi’ao Island, Zhujiang River estuary [J]. Acta Oceanologica Sinica, 2018, 40(7): 79-89. [19] 雷振胜, 李玫, 廖宝文. 珠海淇澳红树林湿地生物多样性现状及保护[J]. 广东林业科技, 2008, 24(5):56-60. [LEI Zhengsheng, LI Mei, LIAO Baowen. Current situation and protection of mangrove wetlands biodiversity on Qi'ao Island, Zhuhai city [J]. Guangdong Forestry Science and Technology, 2008, 24(5): 56-60. [20] 刘俊勇, 林凤标, 刘壮添, 等. 伶仃洋河口近年潮位变化分析及原因探讨[J]. 人民珠江, 2012, 33(6):14-18. doi: 10.3969/j.issn.1001-9235.2012.06.004 [21] 肖志建, 李团结, 廖世智. 伶仃洋表层沉积物特征及其泥沙运移趋势[J]. 热带海洋学报, 2011, 30(4):58-65. [XIAO Zhijian, LI Tuanjie, LIAO Shizhi. Surface sediment characteristics and transport trend in Lingdingyang Bay of the Pearl River estuary [J]. Journal of Tropical Oceanography, 2011, 30(4): 58-65. doi: 10.3969/j.issn.1009-5470.2011.04.009 [22] 丁芮. 珠江口及邻近海域环流潮汐高分辨率数值模拟研究[D]. 中国海洋大学硕士学位论文, 2015. DING Rui. Three-Dimensional high-resolution numercal study of the tide and circulation in the Pearl River estuary and its adjacent wayers[D]. Master Dissertation of Ocean University of China, 2015.
[23] 韩玉梅, 包芸, 任杰. 聚类分析方法在泥沙粒径分级模拟中的应用研究[J]. 烟台大学学报: 自然科学与工程版, 2006, 19(4):288-294. [HAN Yumei, BAO Yun, REN Jie. Application of clustering analysis to simulation model of different sediment grain size [J]. Journal of Yantai University: Natural Science and Engineering Edition, 2006, 19(4): 288-294. [24] 王树功, 黎夏, 刘凯, 等. 近20年来淇澳岛红树林湿地景观格局分析[J]. 地理与地理信息科学, 2005, 21(2):53-57. [WANG Shugong, LI Xia, LIU Kai, et al. Landscape pattern analyses on the mangrove forest wetland of Qi'ao island in the last two decades [J]. Geography and Geo-Information Science, 2005, 21(2): 53-57. doi: 10.3969/j.issn.1672-0504.2005.02.013 [25] 张乔民, 于红兵, 陈欣树, 等. 红树林生长带与潮汐水位关系的研究[J]. 生态学报, 1997, 17(3):258-265. [ZHANG Qiaomin, YU Hongbing, CHEN Xinshu, et al. The relationship between mangrove zone on tidal flats and tidal levels [J]. Acta Ecologica Sinica, 1997, 17(3): 258-265. doi: 10.3321/j.issn:1000-0933.1997.03.006 [26] 王树功, 黎夏, 周永章, 等. 珠江口淇澳岛红树林湿地变化及调控对策研究[J]. 湿地科学, 2005, 3(1): 13-20. WANG Shugong, LI Xia, ZHOU Yongzhang, et al. The change of mangrove wetland ecosystemetland Science, 2005, 3(1): 13-20.
[27] Stuiver M R P J. CALIB 7. 1[WWW program] at http://calib.org, access and controlling countermeasures in the Qi'ao island[J]. Wed 2018-10-8. 2018.
[28] Southon J, Kshgarian M, Fontugne M, et al. Marine reservoir corrections for the Indian Ocean and Southeast Asia [J]. Radiocarbon, 2002, 44(1): 167-180. doi: 10.1017/S0033822200064778
[29] Zong Y Q, Yu F L, Huang G Q, et al. The history of water salinity in the Pearl River estuary, China, during the Late quaternary [J]. Earth Surface Processes and Landforms, 2010, 35(10): 1221-1233. doi: 10.1002/esp.2030
[30] 韩爱艳, 曾砺锋, 黄康有, 等. 罗霄山脉山地沼泽全新世以来的古气候记录[J]. 热带地理, 2016, 36(3):477-485, 520. [HAN Aiyan, ZENG Lifeng, HUANG Kangyou, et al. Holocene climate records from a mountain wetland in Luoxiao ranges [J]. Tropical Geography, 2016, 36(3): 477-485, 520. [31] Blaauw M, Christen J A. Flexible paleoclimate age-depth models using an autoregressive gamma process [J]. Bayesian Analysis, 2011, 6(3): 457-474.
[32] Grimm E C. CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares [J]. Computers & Geosciences, 1987, 13(1): 13-35.
[33] 李淑鸾. 珠江口底质中有孔虫埋葬群的分布规律[J]. 海洋地质与第四纪地质, 1985, 5(2):83-104. [LI Shuluan. Distribution of the foraminiferal thanatocoenosis of Pearl River estuary [J]. Marine Geology & Quaternary Geology, 1985, 5(2): 83-104. [34] 李涛, 向荣, 李团结. 珠江口表层沉积物底栖有孔虫分布及环境指示[J]. 海洋地质与第四纪地质, 2011, 31(6):91-98. [LI Tao, XIANG Rong, LI Tuanjie. Benthic foraminiferal distribution in surface sediments of Zhujiang Estuary and its environmental implications [J]. Marine Geology & Quaternary Geology, 2011, 31(6): 91-98. [35] Wu J, Liu C L, Fürsich F T, et al. Foraminifera as environmental indicators and quantitative salinity reconstructions in the Pearl River Estuary, Southern China [J]. The Journal of Foraminiferal Research, 2015, 45(3): 205-219. doi: 10.2113/gsjfr.45.3.205
[36] 孙息春. 广东深圳湾和海南新村港底质中的有孔虫[J]. 微体古生物学报, 1991, 8(3):325-337. [SUN Xichun. Foraminifera from sediments of the Shenzhen Bay, Guangdong, and the Xincun Harbour, Hainan Island [J]. Acta Micropalaeontologica Sinica, 1991, 8(3): 325-337. [37] Berkeley A, Perry C T, Smithers S G, et al. The spatial and vertical distribution of living (stained) benthic foraminifera from a tropical, intertidal environment, north Queensland, Australia [J]. Marine Micropaleontology, 2008, 69(2): 240-261. doi: 10.1016/j.marmicro.2008.08.002
[38] Edwards R J, Wright A J, Van De Plassche O. Surface distributions of salt-marsh foraminifera from Connecticut, USA: modern analogues for high-resolution sea level studies [J]. Marine Micropaleontology, 2004, 51(1-2): 1-21. doi: 10.1016/j.marmicro.2003.08.002
[39] De Rijk S, Troelstra S R. Salt marsh foraminifera from the Great Marshes, Massachusetts: environmental controls [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1997, 130(1-4): 81-112. doi: 10.1016/S0031-0182(96)00131-9
[40] Patterson R T, Gehrels W R, Belknap D F, et al. The distribution of salt marsh foraminifera at Little Dipper Harbour New Brunswick, Canada: implications for development of widely applicable transfer functions in sea-level research [J]. Quaternary International, 2004, 120(1): 185-194. doi: 10.1016/j.quaint.2004.01.017
[41] Kemp A C, Horton B P, Vann D R, et al. Quantitative vertical zonation of salt-marsh foraminifera for reconstructing former sea level; an example from New Jersey, USA [J]. Quaternary Science Reviews, 2012, 54: 26-39. doi: 10.1016/j.quascirev.2011.09.014
[42] Gehrels W R. Determining relative sea-level change from salt-marsh foraminifera and plant zones on the coast of Maine, U.S.A. [J]. Journal of Coastal Research, 1994, 10(4): 990-1009.
[43] Patterson R T, Dalby A P, Roe H M, et al. Relative utility of foraminifera, diatoms and macrophytes as high resolution indicators of paleo-sea level in coastal British Columbia, Canada [J]. Quaternary Science Reviews, 2005, 24(18-19): 2002-2014. doi: 10.1016/j.quascirev.2004.11.013
[44] Scott D B, Medioli F S. Quantitative Studies of Marsh Foraminiferal Distributions in Nova Scotia: Implications for Sea Level Studies[M]. Washington, D.C.: Cushman Foundation for Foraminiferal Research, 1980.
[45] Sen A, Ghosh M, Khanderao P, et al. Modern benthic foraminiferal assemblages from the world’s largest deltaic mangrove ecosystem, the Sundarbans [J]. Marine Biodiversity, 2015, 46(2): 421-431.
[46] 许艳, 王拓夫. 湛江红树林保护区现代沉积物粒度特征及其对风暴事件的响应[J]. 应用海洋学学报, 2011, 30(2):269-274. [XU Yan, WANG Tuofu. Characteristics of sediment particle size and their response to storm surge in the Zhanjiang Mangrove Nature Reserve [J]. Journal of Oceanography in Taiwan Strait, 2011, 30(2): 269-274. doi: 10.3969/J.ISSN.1000-8160.2011.02.018 [47] 刘金铃, 李柳强, 林慧娜, 等. 中国主要红树林区沉积物粒度分布特征[J]. 厦门大学学报: 自然科学版, 2008, 47(6):891-893. [LIU Jingling, LI Liuqiang, LIN Huina, et al. Characters of Gain Size of Sediments from Mangrove Wetlands of China [J]. Journal of Xiamen University: Natural Science, 2008, 47(6): 891-893. [48] Berkeley A, Perry C T, Smithers S G, et al. A review of the ecological and taphonomic controls on foraminiferal assemblage development in intertidal environments [J]. Earth-Science Reviews, 2007, 83(3-4): 205-230. doi: 10.1016/j.earscirev.2007.04.003
[49] Zong Y. SEA-LEVELS, LATE QUATERNARY | Late quaternary relative sea-level changes in the Tropics[]//Elias S A, Elias C J. Encyclopedia of Quaternary Science. 2nd ed. Oxford, UK: Elsevier, 2013, 4: 495-502.
[50] Xiong H X, Zong Y Q, Qian P, et al. Holocene sea-level history of the northern coast of South China Sea [J]. Quaternary Science Reviews, 2018, 194: 12-26. doi: 10.1016/j.quascirev.2018.06.022
[51] Zong Y Q, Huang K Y, Yu F L, et al. The role of sea-level rise, monsoonal discharge and the palaeo-landscape in the early Holocene evolution of the Pearl River delta, southern China [J]. Quaternary Science Reviews, 2012, 54: 77-88. doi: 10.1016/j.quascirev.2012.01.002
[52] Zong Y Q, Zheng Z, Huang K Y, et al. Changes in sea level, water salinity and wetland habitat linked to the late agricultural development in the Pearl River delta plain of China [J]. Quaternary Science Reviews, 2013, 70: 145-157. doi: 10.1016/j.quascirev.2013.03.020
[53] Zong Y, Yu F, Huang G, et al. Sedimentary evidence of Late Holocene human activity in the Pearl River delta, China [J]. Earth Surface Processes and Landforms, 2010, 35(9): 1095-1102. doi: 10.1002/esp.1970
[54] 王建华, 王晓静, 曹玲珑, 等. 珠江三角洲GZ-2孔全新统孢粉特征及古环境意义[J]. 古地理学报, 2009, 11(6):661-669. [WANG Jianhua, WANG Xiaojing, CAO Linglong, et al. The Holocene sporopollen characteristics and their paleoenvironmental significance of Core GZ-2 in Pearl River delta [J]. Journal of Palaeogeography, 2009, 11(6): 661-669. doi: 10.7605/gdlxb.2009.06.006 [55] 黄康有, 何嘉卉, 宗永强, 等. 珠江三角洲三水盆地早全新世以来孢粉分析与古环境重建[J]. 热带地理, 2016, 36(3):364-373. [HUANG Kangyou, HE Jiahui, ZONG Yongqiang, et al. Holocene paleoenvironment reconstruction based on pollen data in the Sanshui Basin, Northern Pearl River delta [J]. Tropical Geography, 2016, 36(3): 364-373. [56] Berkeley A, Perry C T, Smithers S G, et al. Foraminiferal biofacies across mangrove-mudflat environments at Cocoa Creek, north Queensland, Australia [J]. Marine Geology, 2009, 263(1-4): 64-86. doi: 10.1016/j.margeo.2009.03.019
[57] 余克服, 陈特固. 南海北部晚全新世高海平面及其波动的海滩沉积证据[J]. 地学前缘, 2009, 16(6):138-145. [YU Kefu, CHEN Tegu. Beach sediments from northern South China Sea suggest high and oscillating sea level during the late Holocene [J]. Earth Science Frontiers, 2009, 16(6): 138-145. doi: 10.3321/j.issn:1005-2321.2009.06.015 [58] 李平日. 珠江三角洲一万年来环境演变[M]. 北京: 海洋出版社, 1991: 130-139. LI Pingri. The Environment Evolution of The Zhujiang Delta in The Last 10000 Years[M]. Beijing: China Ocean Press, 1991: 130-139.
[59] 王建华. 华南沿海全新世海滩岩的特征及其意义[J]. 中山大学学报论丛, 1992(1):111-122. [WANG Jianhua. Characteristics and significancos of the Holocene Beachrocks along the coast of South China [J]. Sun Yatsen University Forum, 1992(1): 111-122. [60] 于学峰, 周卫建, 史江峰. 度量泥炭腐殖化度的一种简便方法: 泥炭灰度[J]. 海洋地质与第四纪地质, 2005, 25(1):133-136. [YU Xuefeng, ZHOU Weijian, SHI Jiangfeng. A convenient method for peat humification determination: Grayscale [J]. Marine Geology & Quaternary Geology, 2005, 25(1): 133-136. [61] 马巧红, 钟巍, 薛积彬, 等. 晚更新世晚期以来雷州半岛北部泥炭腐殖化度的古气候意义[J]. 热带地理, 2008, 28(6):498-503. [MA Qiaohong, ZHONG Wei, XIE Jishan, et al. Paleoclimatic significance of the peat humification degree in Northern Leizhou Peninsula since Late-pleistocene [J]. Tropical Geography, 2008, 28(6): 498-503. doi: 10.3969/j.issn.1001-5221.2008.06.002 [62] 王晓静, 王建华, 曹玲珑, 等. 广州地区晚第四纪孢粉气候研究[J]. 中山大学学报: 自然科学版, 2010, 49(3):113-121. [WANG Xiaojing, WANG Jianhua, CAO Linglong, et al. Late Quaternary pollen records and climate significance in Guangzhou [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2010, 49(3): 113-121. [63] 王爱军, 高抒, 陈坚, 等. 福建泉州湾盐沼对台风“格美”的沉积动力响应[J]. 科学通报, 2009, 54(1):120-130. [WANG Aijun, GAO Shu, CHEN Jian, et al. Sediment dynamic responses of coastal salt marsh to typhoon “KAEMI” in Quanzhou Bay, Fujian Province, China [J]. Chinese Science Bulletin, 2009, 54(1): 120-130. doi: 10.1007/s11434-008-0365-7 -
期刊类型引用(3)
1. 刘博华,吴芳,张绪教,崔加伟,董晓朋. 青藏高原东北缘红寺堡盆地晚更新世沉积物元素地球化学特征及其环境指示意义. 地质通报. 2024(01): 33-45 .  百度学术
百度学术
2. 蒋富清,周晓静,李传顺. 220 ka以来西菲律宾海黏土矿物输入变化及其控制因素. 海洋地质与第四纪地质. 2023(05): 85-95 . 本站查看
3. 颜钰,蒋富清,曾志刚. 西太平洋暖池区沉积物源示踪研究进展. 海洋地质与第四纪地质. 2023(05): 148-159 . 本站查看
其他类型引用(2)











计量
- 文章访问数: 2528
- HTML全文浏览量: 452
- PDF下载量: 49
- 被引次数: 5


