Characteristics of the stratigraphic architectures of the shallow sections in deep sea basin on both sides of Kyushu-Palau ridge
-
摘要: 由于欧亚板块、澳大利亚板块与太平洋板块运动的作用,菲律宾海盆形成了全球最为复杂的海底地形地貌。以九州-帕劳海脊为界,西菲律宾海盆与帕里西维拉海盆呈现不同的构造走向与地形特征,海脊两侧发育丰富的深海地质现象与地貌特征。参量阵浅地层剖面仪因其差频窄波束的发射特点,能够获得深海海底浅地层的高分辨率剖面。本文利用参量阵浅地层剖面仪在九州-帕劳海脊获取的高分辨率浅地层剖面,分析了横跨海脊东西走向的海底滑坡发育特点、丘状起伏沉积差异、浅层气以及海底流体运移等地质特征,并结合构造与地形地貌特点对上述特征的成因机制进行了探讨,在垂向小时间尺度上完成了对研究区浅部地层结构的特征研究,为该区域的沉积学和地质学研究提供了思路。Abstract: The Philippine Basin has the most complex submarine topography in the world due to the movement of the three plates, the Eurasian the Australian and the Pacific. Taking the Kyushu-Palau ridge as a boundary, the West Philippine Basin shows different structural patterns and topographic features with the Parece Vela Basin where abundant deep-sea geological phenomena and geomorphic features occur.. The high-resolution profiles of the deep seabed can be acquired with the parametric array shallow layer profiler forits transmitting characteristics of difference-frequency and narrowband beam. In this paper, the characteristics of submarine landslides, the sedimentary differences around sea-knolls and the migration of shallow gas and submarine fluid are carefully studied with the East-West trending shallow strata sections across the ridge. The Genetic mechanism of the geological characteristics is discussed by the integration of structural and topographical features. The research of shallow stratigraphic architectures made on vertical short-time scale may provide more evidence and ideas for the sedimentological and geological researches of the region.
-
菲律宾海板块经过晚中生代时期的三大板块俯冲、碰撞,在弧后扩张、走滑断裂等地质构造作用下,形成了复杂的海底地形地貌特征[1-5]。形成于45 Ma以前的西菲律宾海盆是菲律宾海中最大的盆地,由于四周复杂地形地貌环绕,使其陆源沉积汇入量与海底沉积扰动较少,较为原始的沉积地貌特征得以保存[6-7]。深海沉积特征对于沉积物源的识别与沉积动力过程的研究[8-9]、海底滑坡灾害[10]以及古气候旋回关系[11]等方面的研究有着重要意义。近南北向伸展的九州-帕劳海脊是西菲律宾海盆的东侧边界,并与中央海盆裂谷同为沟弧盆系统的古构造。九州-帕劳海脊的裂解与帕里西维拉海盆的弧后拉张转换约发生在25 Ma,剧烈的构造运动使得火山物质成为主要沉积物源,而自25 Ma以来,该地区接受稳定沉积[12]。前人曾利用地球物理手段获得大型地震反射剖面来研究深层地质构造[13],利用表层地质取样获得表层沉积物的地球化学特征[14],但对该研究区域的高分辨浅地层剖面的研究较为缺乏,无法获得垂向小时间尺度的沉积特征。
参量阵浅地层剖面探测作为当今海洋浅地层调查的重要手段,利用声波在介质中传播的非线性特征,实现差频窄波束声波发射,从而获得高分辨率的地震剖面[15]。参量阵浅地层剖面探测仪不仅有助于查明海底管线布设、海底浅部地层沉积和海底滑坡等地质情况,而且可对浅层气的近海底声学反射特征的探明提供帮助[16]。本文利用跨越九州-帕劳海脊的深海高分辨率浅地层剖面,结合多波束测深数据,对该区域的海底浅部地层剖面特征进行分析与描述,并探讨高分辨浅地层剖面的研究意义。
1. 区域地质背景
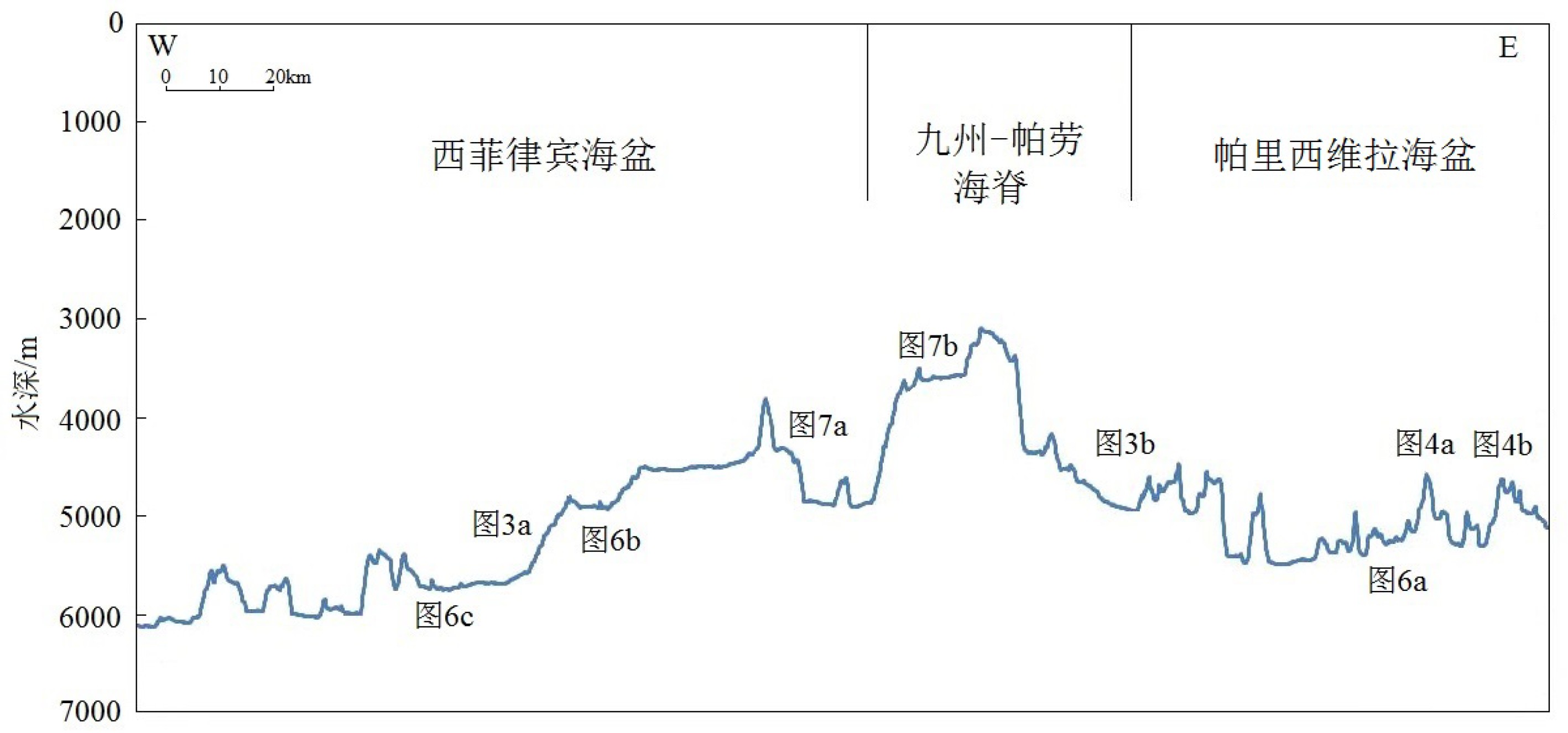
菲律宾海板块位于欧亚板块、澳大利亚板块与太平洋板块之间(图1),受到三大板块运动俯冲与碰撞的影响,板块内发育丰富而复杂的构造单元,例如深海盆地、扩张中心、火山弧以及深海海脊等。九州-帕劳海脊位于菲律宾海盆中部,长度约2750 km,水深范围为3700~5000 m,贯穿南北分隔开西菲律宾海盆与东侧的四国-帕里西维拉海盆[17]。由于太平洋板块西向俯冲菲律宾海板块,强烈的火山活动形成了伊豆-小笠原岛弧古构造[18-20],根据钻探岩心沉积物的Ar元素测年,伊豆-小笠原-马里亚纳岛弧与九州-帕劳海脊均形成于47~45 Ma[21],西菲律宾海盆中央裂谷在54~33 Ma期间,东西两侧进行了不对称的扩张运动[22-23]。29 Ma左右太平洋板块西向俯冲速率降低,西菲律宾海盆的扩张活动也已停止,受到东向拉伸力的作用,伊豆-小笠原-马里亚纳海沟开始向东后退,南端的帕里西维拉海盆最先开始裂开,逐步向北延伸至四国海盆,15 Ma帕里西维拉海盆与四国海盆扩张活动结束,老的伊豆-小笠原岛弧裂解完成,九州-帕劳海脊残留弧结构形成。因此,九州-帕劳海脊东西两侧是不同的扩张与拉伸作用形成,导致两侧坡度[24]、水深[25]、沉积环境[26-27]等均有差异。九州-帕劳海脊两侧的复杂构造背景,使得该区域有较高的研究价值[28-29]。
![图 1 菲律宾海板块及邻区主要构造单元图[13]]()
2. 数据获取与处理
本文浅地层剖面数据采用全海深参量阵浅层剖面仪Parasound P70,浅剖换能器固定安装于调查船底,采集过程中能够保持较好姿态。本文应用测线由东向西横跨九州-帕劳海脊,两侧地形地貌表现各异,浅部地层反射信息丰富。当参量阵浅地层剖面调查同步多波束采集,参量阵的和频与多波束的主频相近时,会相互造成干扰,因此本文选择差频为3.5 kHz的浅地层剖面,波束发射间距根据水深值自适应调节,以满足不同水深变化的调查工作。浅剖数据处理过程较为简单,经过波束触发时间校正后,利用带通滤波处理涌浪噪声与环境噪声,衰减随机干扰噪音,获得反射特征清晰、高信噪比的浅地层剖面。
3. 研究区域浅地层剖面特征与成因分析
本文应用测线长度374.6 km,自东向西依次经过西菲律宾海盆东侧、九州-帕劳海脊以及帕里西维拉海盆西侧,水深值变化剧烈且高差达到3030 m(图2)。
3.1 海底滑坡
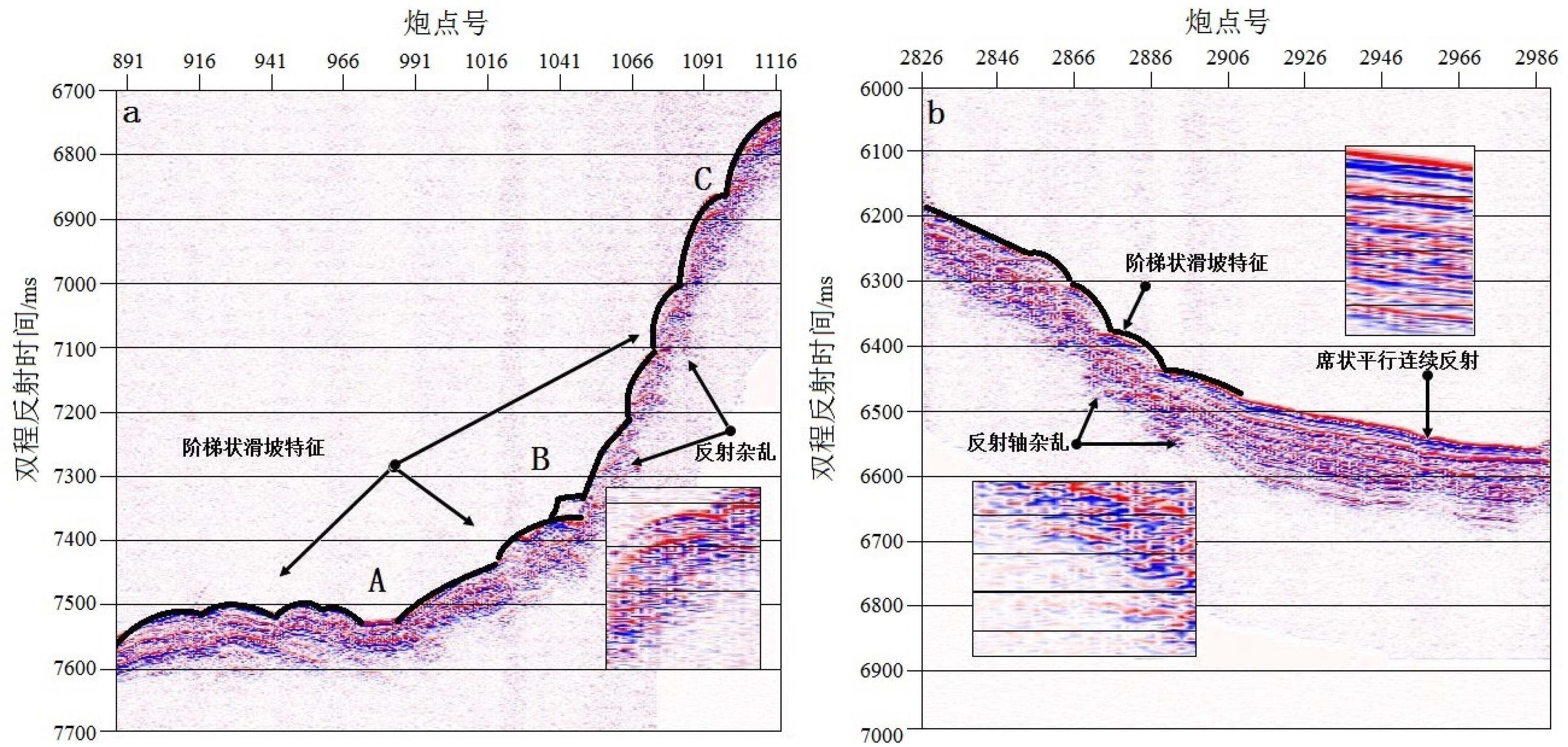
海底滑坡是指海底沉积物受到地形、构造条件、水动力以及物源类型等各种因素影响下的一种运动方式[30],也是海底沉积物搬运的方式之一。海底滑坡因其发生频率高、破坏性大成为近20年海洋研究热点,其形成环境、发生机制以及运动方式的复杂性,导致海底滑坡的分类一直没有较好的共识[31-32]。但是从广义的概念来讲,海底滑坡的过程就是海底沉积物的搬运过程,包括蠕动、崩塌以及重力流现象[33]。目前常用地球物理手段间接地观察海底沉积物边界特征,推断出海底滑坡的运动过程。大陆边缘的河口三角洲、深海沉积扇、深海陆坡以及海脊均是海底滑坡容易发生的区域,分析其原因在于海底滑坡的发生机制需要一定量的沉积物、一定的搬运距离、沉积物的抵抗剪切强度以及内在的力学驱动。从高分辨浅地层剖面能够分析出海脊两侧呈现因剧烈构造运动而发生沉积物滑坡的反射特征(图3)。图3a最大坡度为4.76°,而图3b最大坡度为1.77°,两剖面具有不同的倾斜角度、不同的滑坡距离以及沉积物再沉积的厚度。
![图 3 海底滑坡高分辨浅地层剖面图]() 图 3 海底滑坡高分辨浅地层剖面图Figure 3. High resolution shallow stratigraphic profile of submarine landslide
图 3 海底滑坡高分辨浅地层剖面图Figure 3. High resolution shallow stratigraphic profile of submarine landslide通常深海峡谷具有一定的坡度接受原始的沉积物积累,并且坡上的重力分量能够提供海底滑坡的内在力学驱动,底部较为平坦的地势为沉积物提供了滑塌所需要的搬运距离,深海沉积物含水量较大,造成沉积物的低抗剪切强度。沉积物滑坡过程与地层抬升过程相对应,根据地形坡度可以看出图3a中经历两次不同程度的地层抬升。AB段坡度为1.07°,沉积物在某个角度以较小的初始速度开始滑坡,滑坡量不大,缓慢堆积于坡底。随着沉积活动与抬升活动的继续发生,BC段抬升动力更大,获得更大的坡度(约4.08°),沉积物滑坡速度较快,并且在滑塌的过程中挤压推动坡底的沉积物,使其在平坦的坡底进行再沉积,沉积物同相轴连续清晰表明再沉积速度较慢,时间较长。AB段由于上覆BC段的沉积物推动,反射同相轴出现错断不连续。BC段由于滑坡速度较快的原因,同相轴杂乱不连续。图3b中,由于整个剖面坡度只有1.77°,因此沉积物获得加速度较小,搬运过程较弱,整个过程均发生在坡上,并且根据坡上的反射轴特征,该区域沉积物搬运过程较图3a缓慢,底部呈现席状平行的反射特征。
该区域的滑坡特征说明,海底地形坡度提供的内在驱动力是海底沉积物滑坡的影响因素,即使在小坡度的情况下仍然能够诱发缓慢的沉积物滑动[34-35],因此,海底地形的坡度能够影响沉积物滑坡过程中的再沉积速度与沉积特征。
3.2 丘状起伏沉积差异
海脊东侧区域处于帕里西维拉海盆中,受到东向拉伸力的作用,该区域由一系列复杂的丘状起伏组成,地形走向近南北方向[25]。帕里西维拉海盆扩张过程中,西部沉积物厚度较小,且以远洋黏土沉积为主[36]。深海沉积物的来源中,陆源比例很少,主要为海洋生物源、海底火山物源等。影响沉积物分布的因素有地形地貌、水动力、气候变化以及海平面涨落。其中洋流、底流以及海底构造等活动使得海洋环境处于动态平衡状态中,沉积物在动态的海洋环境中会发生再次搬运与沉积[37]。分析高分辨率浅地层剖面发现,不同地形结构的相邻区域存在沉积物厚度差异(图4)。图4a与图4b中同一区域的反射特征均呈现丘状起伏、狭窄深谷处的沉积层明显更薄,沉积厚度最大相差近40 m。
![图 4 海脊东侧丘状起伏地形沉积差异]() 图 4 海脊东侧丘状起伏地形沉积差异图c中黄色线为施工测线,矩形范围对应图a与图b位置。Figure 4. differences in deposition around the hilly terrain on the east side of the ridgeThe yellow shows is the survey line. The rectangular range corresponds to position a and b in c.
图 4 海脊东侧丘状起伏地形沉积差异图c中黄色线为施工测线,矩形范围对应图a与图b位置。Figure 4. differences in deposition around the hilly terrain on the east side of the ridgeThe yellow shows is the survey line. The rectangular range corresponds to position a and b in c.海脊东侧的帕里西维拉海盆内部存在的底流来自于南极底层水[38]。受到该区域地形的影响,底流会绕过障碍物流动,围绕整个海盆呈现逆时针转动(图5),在多项相关研究中,该区域的底层流速度较快[39],平均速度达到15 cm/s。当海底层流经过丘状起伏地形,流体速度因流体横截面积的影响会发生局部改变。在流体通量一定的情况下,流体速度与流经的横截面积成反比。在图4中a与b两处的窄沟跨度均小于1 km,与宽度达到2 km的宽沟相比,流速提高一倍。深海沉积以远洋沉积物源为主,流速的差异造成沉积速率与沉积物搬运速度的不同,进而形成了不同间距的丘状起伏处的沉积物厚度差异。
![图 5 帕里西维拉海盆底层流向示意图[39]]()
3.3 浅层气与海底流体运移
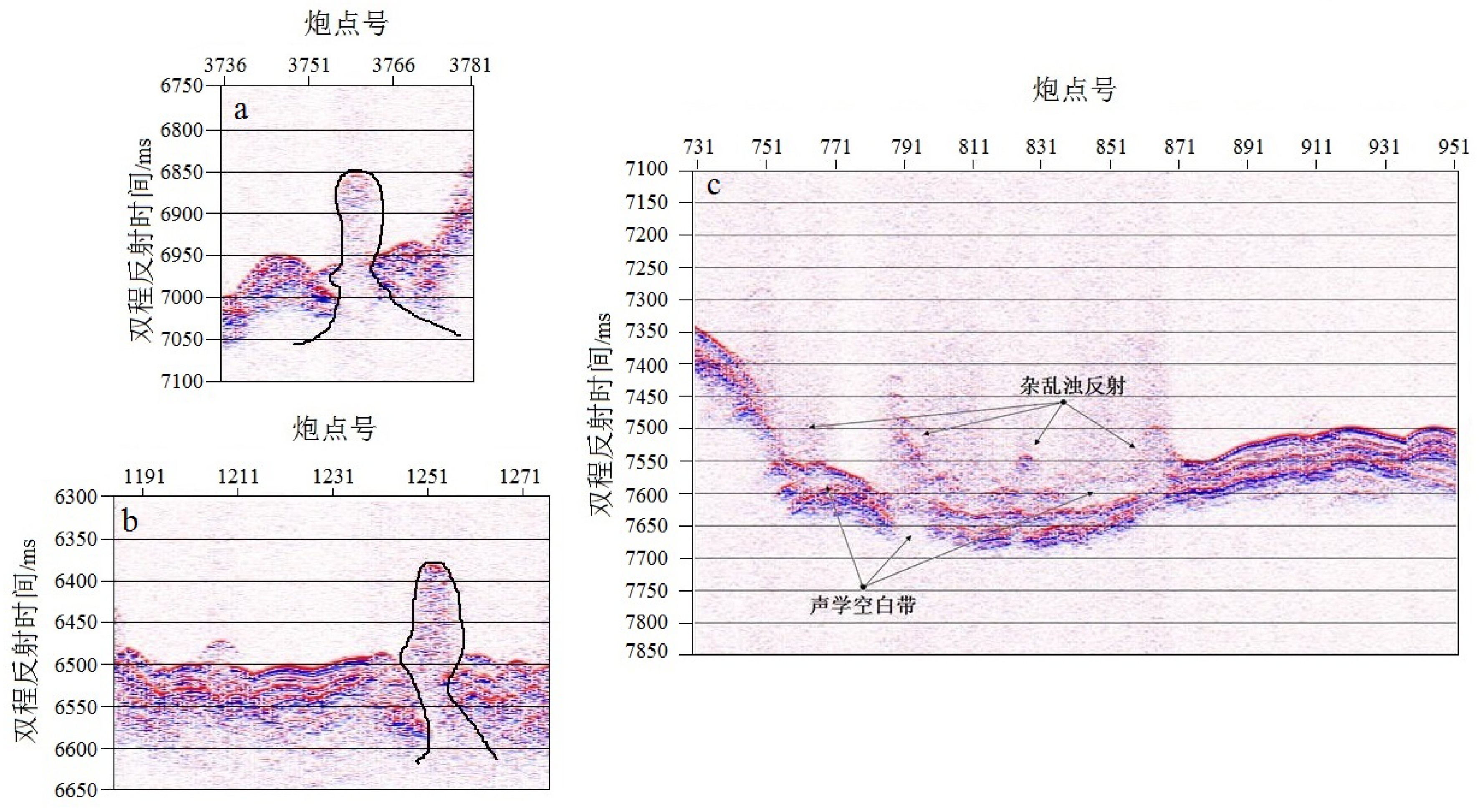
深海浅层气渗漏、冷泉以及热液活动等现象是当今深海大洋研究的热点,这些现象常常与海底资源相关,冷泉与热液活动的极端系统的相关性研究,能够揭示岩石圈与外部圈层的物质、能量的交换与转移[39]。浅层气的存在能够引起沉积层的区域地质条件的不稳定,对人类生活与海洋工程等造成影响[40]。由于存在声阻抗差,该类活动在反射剖面上具有明显的特征,例如浊反射、声学空白带、云状声反射以及羽状流等,并常出现泥火山、麻坑、自生性碳酸盐岩等海底地质特征[41]。
分析高分辨浅地层剖面可以发现,在海脊两侧的深海盆地海底浅层气发育丰富。由于远洋沉积厚度较薄且沉积时间长,浅层气较容易渗漏出海底并干扰沉积物成像(图6)。声图呈“云状”反射且浅部地层出现声反射空白区,沉积物反射轴杂乱不连续,浅层气通道边界明显。当浅层气渗漏面积较大且地形起伏较快时,参量阵高分辨浅地层剖面受到干扰更为严重(图6c)。由于浅层气侵入表层沉积物,造成其反射轴连续性变差,形成明显的声学空白区,以测深数据为约束,对比水深值与地形起伏情况,可分析固液界面的杂乱浊反射范围。浅层气渗漏以及海底流体超压上涌形成的麻坑与泥火山,结合多波束测深数据确定位置信息,能够确认其浅地层剖面特征(图7)。
![图 6 浅层气渗漏及其浊反射剖面特征]() 图 6 浅层气渗漏及其浊反射剖面特征Figure 6. Shallow gas leakage and its characteristics of shallow gas turbidity reflection profile
图 6 浅层气渗漏及其浊反射剖面特征Figure 6. Shallow gas leakage and its characteristics of shallow gas turbidity reflection profile![图 7 麻坑及泥火山]() 图 7 麻坑及泥火山c与d为对应多波束资料。Figure 7. Pockmark and mud volcanoc and d correspond to multi-beam data.
图 7 麻坑及泥火山c与d为对应多波束资料。Figure 7. Pockmark and mud volcanoc and d correspond to multi-beam data.相比近陆地沉积,深海沉积物源相对单一,深海浅层气主要是来自于热成甲烷气[42]。热成甲烷气通常是在高温高压的深地层中产生,形成了深海浅层气的气源。研究区域两侧的深海海盆,均存在中央裂谷拉伸活动的扩张,盆地中发育丰富的断层与裂隙[2-7],为深层甲烷气提供了上升通道。甲烷气通过运移通道聚集于浅地层中,形成海底浅层气。当沉积层受到水动力、地质构造等作用的扰动,浅层气易于渗漏出海底。不同压力值使得浅层气呈现不同的反射特征,会在浅地层反射剖面上出现羽状流、声学空白区域以及入侵表层沉积物引起的反射轴错断。
4. 结论
(1)九州-帕劳海脊两侧地质现象的多样性来自于构造作用下地形地貌的复杂性。该区域发育有丰富的海底滑坡、丘状起伏地形沉积差异、浅层气聚集与海底流体运移等地质现象。浅地层剖面上滑坡距离与再沉积速度等特征,揭示了滑坡特征与坡度的关系。水动力等环境因素直接制约了深海沉积物的沉积特征,造成了丘状起伏地形的沉积厚度差异。深海海盆扩张与海脊快速隆起的构造作用,为圈层物质交换提供了可能性,使得热成气聚集于浅层沉积物中甚至渗漏出海底,海底流体受到超压作用影响向上运移,在浅层气共同的作用下刺穿海底,形成泥火山与麻坑。
(2)参量阵浅地层剖面仪能够对海底几百米以内的沉积地层形成高分辨图像,应用于深海浅层沉积物的形成、运移等方面的研究。结合多波束测深、多道地震等数据,构建深海地层垂直方向表—浅—深的全尺度海底地层体系,有利于开展深海地形地貌成像、沉积特征以及板块构造作用的系统性研究。
致谢:本次研究使用的数据由青岛海洋地质研究所“海洋地质九号”船深海调查团队获取,数据处理中心团队进行室内处理,在此表示感谢。
-
![]()
![]()
图 3 海底滑坡高分辨浅地层剖面图
Figure 3. High resolution shallow stratigraphic profile of submarine landslide
![]()
图 4 海脊东侧丘状起伏地形沉积差异
图c中黄色线为施工测线,矩形范围对应图a与图b位置。
Figure 4. differences in deposition around the hilly terrain on the east side of the ridge
The yellow shows is the survey line. The rectangular range corresponds to position a and b in c.
![]()
![]()
图 6 浅层气渗漏及其浊反射剖面特征
Figure 6. Shallow gas leakage and its characteristics of shallow gas turbidity reflection profile
-
[1] 秦蕴珊, 尹宏. 西太平洋: 我国深海科学研究的优先战略选区[J]. 地球科学进展, 2011, 26(3):245-248. [QIN Yunshan, YIN Hong. Western Pacific: the strategic priority in China deep-Sea research [J]. Advances in Earth Science, 2011, 26(3): 245-248. [2] 李三忠, 余珊, 赵淑娟, 等. 东亚大陆边缘的板块重建与构造转换[J]. 海洋地质与第四纪地质, 2013, 33(3):65-94. [LI Sanzhong, YU Shan, ZHAO Shujuan, et al. Tectonic transition and plate reconstructions of the East Asian continental margin [J]. Marine Geology & Quaternary Geology, 2013, 33(3): 65-94. [3] 王霄飞, 余珊, 李三忠, 等. 板缘与板内活动构造与地震触发机制对比[J]. 海洋地质与第四纪地质, 2014, 34(2):159-175. [WANG Xiaofei, YU Shan, LI Sanzhong, et al. Comparison of active tectonics and earthquakes triggering mechanism along plate boundary and within intraplate [J]. Marine Geology & Quaternary Geology, 2014, 34(2): 159-175. [4] Xu S M, Ye Q, Li S Z, et al. Sequential patterns in Cenozoic marginal basins of the Northwest Pacific [J]. Geological Journal, 2016, 51(S1): 387-415.
[5] Zhang Y, Li S Z, Suo Y H, et al. Origin of transform faults in back-arc basins: examples from Western Pacific marginal seas [J]. Geological Journal, 2016, 51(S1): 490-512.
[6] Hilde T W C, Lee C S. Origin and evolution of the West Philippine Basin: A new interpretation [J]. Tectonophysics, 1984, 102(1-4): 85-104. doi: 10.1016/0040-1951(84)90009-X
[7] 张斌, 李广雪, 黄继峰. 菲律宾海构造地貌特征[J]. 海洋地质与第四纪地质, 2014, 34(2):79-88. [ZHANG Bin, LI Guangxue, HUANG Jifeng. The tectonic geomorphology of the Philippine sea [J]. Marine Geology & Quaternary Geology, 2014, 34(2): 79-88. [8] 李梦君, 毕乃双, 胡丽沙, 等. 南海北部台湾峡谷“蛟龙号”第140潜次沉积物特征及其沉积过程指示意义[J]. 海洋地质与第四纪地质, 2019, 39(4):23-33. [LI Mengjun, BI Naishang, HU Lisha, et al. Sedimentary characteristics and processes revealed by the push cores of the 140th dive of DSV “Jiaolong” in the Taiwan Submarine Canyon, Northern South China Sea [J]. Marine Geology & Quaternary Geology, 2019, 39(4): 23-33. [9] 鄢全树, 石学法, 王昆山, 等. 西菲律宾海盆表层沉积物中的轻碎屑分区及物质来源[J]. 地质评论, 2007, 53(6):765-773. [YAN Quanshu, SHI Xuefa, WANG Kunshan, et al. Provinces and material provenance of light detritus in the surficial sediments from the western Philippine Sea [J]. Geological Review, 2007, 53(6): 765-773. [10] 陈晓辉, 张训华, 李日辉, 等. 渤海海峡海域灾害地质研究[J]. 海洋地质与四纪地质, 2014, 34(1):11-19. [CHEN Xiaohui, ZHANG Xunhua, LI Rihui, et al. A preliminary study on hazardous geology in the Bohai Strait [J]. Marine Geology & Quaternary Geology, 2014, 34(1): 11-19. [11] 段毅, 罗斌杰. 深海现代沉积有机质碳同位素组成变化的古气候证据[J]. 海洋地质与第四纪地质, 1998, 18(4):53-58. [DUAN Yi, LUO Binjie. Palaeoclimate evidence for carbon isotopic changes of organic matters in modern Deep-Sea sediments [J]. Marine Geology & Quaternary Geology, 1998, 18(4): 53-58. [12] 董冬冬, 张正一, 张广旭, 等. 西菲律宾海盆的构造沉积特征及对海盆演化的指示: 来自地球物理大断面的证据[J]. 海洋与湖沼, 2017, 48(6):1415-1425. [DONG Dongdong, ZHANG Zhengyi, ZHANG Guangxu, et al. Tectonic and sedimentary features of the West Philippine Basin and its implication to the basin evolution—evidence from a seismic transection [J]. Oceanologia et Limnologia Sinica, 2017, 48(6): 1415-1425. doi: 10.11693/hyhz20170700178 [13] 丁巍伟, 李家彪. 九州-帕劳海脊南段的深部结构探测及对板块俯冲起始机制的可能启示[J]. 海洋地质与第四纪地质, 2019, 39(5):98-103. [DING Weiwei, LI Jiabiao. Seismic detection of deep structure for Southern Kyueshu-Palau Ridge and its possible implications for subduction initiation [J]. Marine Geology & Quaternary Geology, 2019, 39(5): 98-103. [14] 石学法, 陈丽蓉, 李坤业, 等. 西菲律宾海西部海域粘土沉积物的成因矿物学研究[J]. 海洋地质与第四纪地质, 1995, 15(2):61-72. [SHI Xuefa, CHEN Lirong, LI Kunye, et al. Study on minerageny of the clay sediment in the west of Philippine Sea [J]. Marine Geology & Quaternary Geology, 1995, 15(2): 61-72. [15] 冯京, 尉佳, 刘长春, 等. 参量阵浅剖及电火花震源浅剖互补性分析[J]. 海洋地质前沿, 2019, 35(9):63-68. [FENG Jing, WEI Jia, LIU Changchun, et al. Complementarity analysis of parametric array and spark source sub-bottom profilers [J]. Marine Geology Frontiers, 2019, 35(9): 63-68. [16] 赵铁虎, 张训华, 冯京. 海底油气渗漏浅表层声学探测技术[J]. 海洋地质与第四纪地质, 2010, 30(6):149-156. [ZHAO Tiehu, ZHANG Xunhua, FENG Jing. Acoustic detection techniques for seabed hydrocarbon seepage [J]. Marine Geology & Quaternary Geology, 2010, 30(6): 149-156. [17] Ark J O, Hori T, Kaneda Y. Seismotectonic Implications of the Kyushu-Palau Ridge Subducting Beneath the Westernmost Nankai Forearc [J]. Earth, Planets and Space, 2009, 61(8): 1013-1018. doi: 10.1186/BF03352951
[18] Haraguchi S, Ishii T, Kimura J I. Early tholeiitic and calc-alkaline arc magmatism of middle to late eocene age in the southern Ogasawara (Bonin) forearc [J]. Contributions to Mineralogy and Petrology, 2008, 156(2): 269-272. doi: 10.1007/s00410-008-0307-z
[19] Okino K, Kasuga S, Ohara Y. A new scenario of the parece vela basin genesis [J]. Marine Geophysical Researches, 1998, 20(1): 21-40. doi: 10.1023/A:1004377422118
[20] Hickey-Vargas R. Basalt and Tonalite from the Amami plateau, northern west Philippine basin: new early cretaceous ages and geo-chemical results, and their petrologic and tectonic implications [J]. The Island Arc, 2005, 14(4): 653-665. doi: 10.1111/j.1440-1738.2005.00474.x
[21] Cosca M A, Arculus R J, Pearce J A, et al. 40Ar/39Ar and K-Ar geochronological age constraints for the inception and early evolution of the Izu-Bonin-Mariana arc system [J]. The Island Arc, 1998, 7(3): 579-595. doi: 10.1111/j.1440-1738.1998.00211.x
[22] Deschamps A, Okino K, Fujioka K. Late amagmatic extension along the central and eastern segments of the west Philippine basin fossil spreading axis [J]. Earth and Planetary Science Letters, 2002, 203(1): 277-293. doi: 10.1016/S0012-821X(02)00855-5
[23] Okino K, Fujioka K. The central basin spreading center in the Philippine sea: structure of an extinct spreading center and implications for marginal basin formation [J]. Journal of Geophysical Research: Solid Earth, 2003, 108(B1): 2040.
[24] Kobayashi K. Origin of the Palau and Yap trench-arc systems [J]. Geophysical Journal International, 2004, 157(3): 1303-1315. doi: 10.1111/j.1365-246X.2003.02244.x
[25] 李常珍, 李乃胜, 林美华. 菲律宾海的地势特征[J]. 海洋科学, 2000, 24(6):47-51. [LI Changzhen, LI Naisheng, LIN Meihua. Terrain features of the Philippine sea [J]. Marine Sciences, 2000, 24(6): 47-51. doi: 10.3969/j.issn.1000-3096.2000.06.014 [26] 殷征欣, 李正元, 沈泽中, 等. 西太平洋帕里西维拉海盆不对称性发育特征及其成因[J]. 吉林大学学报: 地球科学版, 2019, 49(1):218-229. [YIN Zhengxin, LI Zhengyuan, SHEN Zezhong, et al. Asymmetric geological developments and their geneses of the Parece Vela basin in western pacific ocean [J]. Journal of Jilin University: Earth Science Edition, 2019, 49(1): 218-229. [27] 林震, 易亮, 王海峰, 等. 西北太平洋采薇海山表层沉积的岩石磁学特征及其对深海环境过程的指示[J]. 地球物理学报, 2019, 62(8):3067-3077. [LIN Zhen, YI Liang, WANG Haifeng, et al. Rock magnetism of deep-sea sediments at Caiwei Guyot, Magellan seamounts of Northwest Pacific and its significance to abyssal environmental changes [J]. Chinese Journal of Geophysics (in Chinese), 2019, 62(8): 3067-3077. doi: 10.6038/cjg2019M0526 [28] 张洁, 李家彪, 丁巍伟. 九州-帕劳海脊地壳结构及其形成演化的研究综述[J]. 海洋科学进展, 2012, 30(4):595-607. [ZHANG Jie, LI Jiabiao, DING Weiwei. Reviews of the study on crustal structure and evolution of the Kyushu-Palau ridge [J]. Advances in Marine Science, 2012, 30(4): 595-607. doi: 10.3969/j.issn.1671-6647.2012.04.016 [29] Ishizuka O, Tani K, Reagan M K, et al. The timescales of subduction initiation and subsequent evolution of an oceanic island arc [J]. Earth and Planetary Science Letters, 2011, 306(3-4): 229-240. doi: 10.1016/j.jpgl.2011.04.006
[30] 李广雪, 庄克琳, 姜玉池. 黄河三角洲沉积体的工程不稳定性[J]. 海洋地质与第四纪地质, 2000, 20(2):21-26. [LI Guangxue, ZHUANG Kelin, JIANG Yuchi. Engineering instability of the deposition bodies in the Yellow River delta [J]. Marine Geology & Quaternary Geology, 2000, 20(2): 21-26. [31] 王大伟, 吴时国, 秦志亮, 等. 南海陆坡大型块体搬运体系的结构与识别特征[J]. 海洋地质与第四纪地质, 2009, 29(5):65-72. [WANG Dawei, WU Shiguo, QIN Zhiliang, et al. Architecture and identification of large Quaternary mass transport depositions in the slope of South China Sea [J]. Marine Geology & Quaternary Geology, 2009, 29(5): 65-72. [32] Prior D B, Coleman J M. Submarine landslides—geometry and nomenclature [J]. Zeitschrift fur Geomorphologie, 1982, 4: 21-33.
[33] 朱超祁, 贾永刚, 刘晓磊, 等. 海底滑坡分类及成因机制研究进展[J]. 海洋地质与第四纪地质, 2015, 35(6):153-163. [ZHU Chaoqi, JIA Yonggang, LIU Xiaolei, et al. Classification and genetic machanism of submarine landslide: a review [J]. Marine Geology & Quaternary Geology, 2015, 35(6): 153-163. [34] Pinet N, Brake V, Campbell C, et al. Geomorphological characteristics and variability of Holocene mass-transport complexes, St. Lawrence River Estuary, Canada [J]. Geomorphology, 2015, 228: 286-302. doi: 10.1016/j.geomorph.2014.09.008
[35] Herzer R H. Uneven submarine topography south of Mernoo Gap—the result of volcanism and submarine sliding [J]. New Zealand Journal of Geology and Geophysics, 1975, 18(1): 183-188. doi: 10.1080/00288306.1975.10426354
[36] Scott R, Kroenke L. Evolution of back arc spreading and arc volcanism in the Philippine Sea: Interpretation of Leg 59 DSDP results[M]//Hayes D E. The Tectonic and Geologic Evolution of Southeast Asian Seas and Islands. Washington, DC: Geophysical Monograph Series, 1980, 23: 283-291.
[37] 明洁. 东菲律宾海帕里西维拉海盆第四纪沉积特征和物质来源及其古环境意义[D]. 中国科学院研究生院(海洋研究所)博士学位论文, 2013. MING Jie. The characteristics and provenance of the sediment in the Parece Vela Basin since the Quaternary and their environmental implications[D]. Doctor Dissertation of the Institute of Oceanology, Chinese Academy of Sciences (Institute of Oceanology), 2013.
[38] Lee I, Ogawa Y. Bottom-current deposits in the Miocene–Pliocene Misaki Formation, Izu forearc area, Japan [J]. Island Arc, 1998, 7(3): 315-329. doi: 10.1111/j.1440-1738.1998.00192.x
[39] 吴能友, 孙治雷, 卢建国, 等. 冲绳海槽海底冷泉-热液系统相互作用[J]. 海洋地质与第四纪地质, 2019, 39(5):23-35. [WU Nengyou, SUN Zhilei, LU Jianguo, et al. Interaction between seafloor cold seeps and adjacent hydrothermal activities in the Okinawa Trough [J]. Marine Geology & Quaternary Geology, 2019, 39(5): 23-35. [40] Campbell K A. Hydrocarbon seep and hydrothermal vent paleoenvironments and paleontology: Past developments and future research directions [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2006, 232(2-4): 362-407. doi: 10.1016/j.palaeo.2005.06.018
[41] 刘伯然, 宋海斌, 关永贤, 等. 南海东北部陆坡冷泉系统的浅地层剖面特征与分析[J]. 地球物理学报, 2015, 58(1):247-256. [LIU Boran, SONG Haibin, GUAN Yongxian, et al. Characteristics and formation mechanism of cold seep system in the northeastern continental slope of South China Sea from sub-bottom profiler data [J]. Chinese Journal of Geophysics, 2015, 58(1): 247-256. doi: 10.6038/cjg20150122 [42] 蔡秋蓉. 海底浅层气——资源还是灾害?[C]//海洋地质环境与资源学术研讨会论文摘要汇编. 广州: 中国地质学会, 2003: 71. CAI Qiurong. Shallow gas on the sea floor: resources or disasters?[C]//Summary of papers of the Symposium on marine geological environment and resources. Guangzhou: China Geological Association, 2003: 71.
-
期刊类型引用(6)
1. 任江波, 王历星, 刘实佳, 姜玉涵, 于淼, 杨永, 姚会强, 何高文. 菲律宾海铁锰结核和结壳的分布规律和成因机制. 岩石学报. 2025(07)  百度学术
百度学术
2. 方中华,陆凯,杨源,冯京,于得水,秦轲,尉佳. 海洋声学地质调查技术现状与展望. 海洋地质与第四纪地质. 2024(03): 82-89 . 本站查看
3. Feifei Zhang,Dingding Wang,Xiaolin Ji,Fanghui Hou,Yuan Yang,Wanyin Wang. Structural features in the mid-southern section of the Kyushu–Palau Ridge based on satellite altimetry gravity anomaly. Acta Oceanologica Sinica. 2024(04): 50-60 . 必应学术
4. 刘佳,李攀峰,邢磊,胡邦琦,杨慧良,赵京涛. 九州-帕劳海脊南段及邻近海域典型海山形态特征及地质意义. 海洋地质与第四纪地质. 2024(04): 65-77 . 本站查看
5. 刘玉斌,宋永东,张建兴,庄丽华,栾振东,阎军. 黑石岛附近海域地形地貌及表层沉积物分布特征分析. 海洋地质前沿. 2023(08): 60-69 . 百度学术
6. 孙美静,罗伟东,陈泓君,胡小三,黄文凯,杜文波,周娇,韩艳飞,刘杰. 西菲律宾海盆中央裂谷周缘地质灾害特征及诱发因素. 海洋学报. 2022(04): 23-35 . 百度学术
其他类型引用(0)
![图 1 菲律宾海板块及邻区主要构造单元图[13]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2021/1/2020072202-1.jpg)
 下载:
下载:

![图 5 帕里西维拉海盆底层流向示意图[39]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2021/1/2020072202-5.jpg)