
Changes in siliceous paleoproductivity during the early Middle Pleistocene in the Northwest Bay of Bengal
-
摘要: 西北孟加拉湾位于印度夏季风的核心降水区,是研究印度季风活动及其对环境影响的理想地区。通过对西北孟加拉湾马哈纳迪盆地IODP353航次U1446站位岩芯生物硅含量和沉积通量的分析,重建了研究区早更新世末—中更新世早期(920~370 ka)的硅质生物生产力演变历史,探讨了印度夏季风降水和径流活动对研究区古生产力变化的影响机制。结果显示,西北孟加拉湾在920~370 ka期间,硅质生产力总体上表现为中更新世过渡期(Mid-Pleistocene Transition,MPT)较低,MPT之后呈上升的趋势;具有明显的冰期-间冰期旋回性,冰期时下降,间冰期上升,且在MPT之后波动幅度增大。对比该站位已有的δ18Osw和Rb/Ca比值等夏季风降水和径流指标发现,印度夏季风增强时硅质生产力上升,减弱时硅质生产力下降,推测印度夏季风的增强导致孟加拉湾降水以及周边河流径流量剧增,河流提供的营养物质使得西北孟加拉湾表层硅质生物勃发,硅质生产力迅速升高。而印度夏季风减弱时,营养盐的减少抑制了表层浮游硅质生物的繁殖,从而使得硅质生产力下降。U1446站位生物硅通量的频谱和小波分析也显示,研究区的硅质生产力受轨道周期变化的调控。Abstract: The Northwest Bay of Bengal (BoB) is located in the core precipitation zone of the Indian Summer Monsoon (ISM), and an ideal area to study Indian monsoon activity and its impact on the environment. We reconstructed the history of siliceous productivity evolution in the study area during late Early Pleistocene to early Middle Pleistocene (920~370 ka) by analyzing biogenic silica content and sediment fluxes of cores from Site U1446 of IODP Expedition 353 in the Mahanadi Basin, Northwest BoB, and explored the mechanism of ISM precipitation and runoff activities on the paleoproductivity changes in the study area. Results indicate that siliceous productivity reflects a trend from low in the Mid-Pleistocene Transition (MPT) to high after MPT in the northwest BoB during 920~370 ka in an obvious glacial-interglacial cycle. It decreased during the glacial period and increased during the interglacial period, and the fluctuation increased after the MPT. A comparison of existing summer monsoon precipitation and runoff indicators such as δ18Osw and Rb/Ca ratio at this site reveals that siliceous productivity increased (decreased) when the ISM strengthened (weakened). It is hypothesized that the enhanced ISM led to a dramatic increase in runoff from the Indian peninsula and precipitation in the BoB, and the nutrients from rivers led to a proliferation of surface siliceous organisms and a rapid upsurge in siliceous productivity in the northwest BoB. However, the weakened ISM and the reduction of nutrients inhibited the flourishing of planktonic siliceous organisms, which then decreased the surface productivity. Spectral and wavelet analyses of biogenic silica fluxes at Site U1446 also indicate that siliceous productivity was modulated by orbital cycle variations in this study area.
-
在古生产力的研究中,生源碳酸钙[1]、有机碳[2]和生物硅含量[3]等都被用来作为恢复古生产力的指标。其中,生物硅对海洋生态系统中的碳循环具有极其重要的影响,全球超过40%的初级生产力要归功于硅藻[4],它如今占全球开阔大洋及海岸带总浮游植物的90%以上[5],控制了大洋表层水向深部海洋中的碳输出,对大气CO2浓度变化有潜在影响[6]。与钙质骨骼生物相比,它对海洋中CO2的影响更加直接[7]。
西北孟加拉湾位于印度季风活动区,同时也是印度夏季风降水和河流汇入的核心区域。夏季风期间大量的陆地淡水输入使得表层海水盐度下降,导致强烈的表层水体分层[8-9],并影响到营养物质的供应[10],控制着西北孟加拉湾地区的初级生产力变化。该区的表层生产力变化和印度夏季风之间有着密切的关系,因而成为研究印度季风演变和古生产力变化的理想场所。以前的研究认为,印度夏季风期间,强烈的低盐表层水分层现象,阻碍了下部水体中的营养物质进入透光带,因而现代孟加拉湾开放大洋表现为较低的生产力。虽然丰富的河流系统为孟加拉湾供应了大量的营养物质和溶解硅,但它们主要被河口和海岸地区的生物活动所消耗,对开放大洋的生产力贡献不大[10]。近来的研究发现,地史时期,西北孟加拉湾近岸地区在夏季风增强期间虽然钙质生物生产力下降,但是硅质生物生产力却显著上升[11-12]。因而了解该区硅质生物生产力的变化,不仅能够揭示地史时期印度夏季风的活动,而且对认识孟加拉湾地区在全球碳循环过程中的作用也具有重要的意义。
第四纪1.25~0.7 Ma期间发生强振幅和高频率的气候振荡,又称中更新世过渡(Mid-Pleistocene Transition, MPT),其最大的特征是冰期-间冰期旋回从41 ka周期向100 ka周期的转变[13]。在此期间,不论陆地还是大洋环境都发生了重要的变化,西北孟加拉湾的古生产力变化在MPT期间也发生了明显的波动。Lee等[11]对西北孟加拉湾U1445站位的古生产力研究发现,此海区2.3 Ma以来可以分成pre-MPT(MPT之前)、MPT和post-MPT(MPT之后)3个阶段,从老到新硅质生物生产力随着季风导致的降水量和海水分层强度的下降而减弱,但由于其分辨率较低,难以对此海区在轨道尺度上的古生产力变化规律及其机制进行探究。
本次研究以国际大洋发现计划(International Ocean Discovery Program, IODP)353航次在西北孟加拉湾马哈纳迪盆地钻取的U1446站位(图1)为研究对象,通过对该钻孔岩芯沉积物样品中生物硅含量的分析,重建西北孟加拉湾跨越中更新世过渡期的表层古生产力演变历史,探讨MPT期及其之后印度夏季风活动对研究区古生产力变化的控制机制。
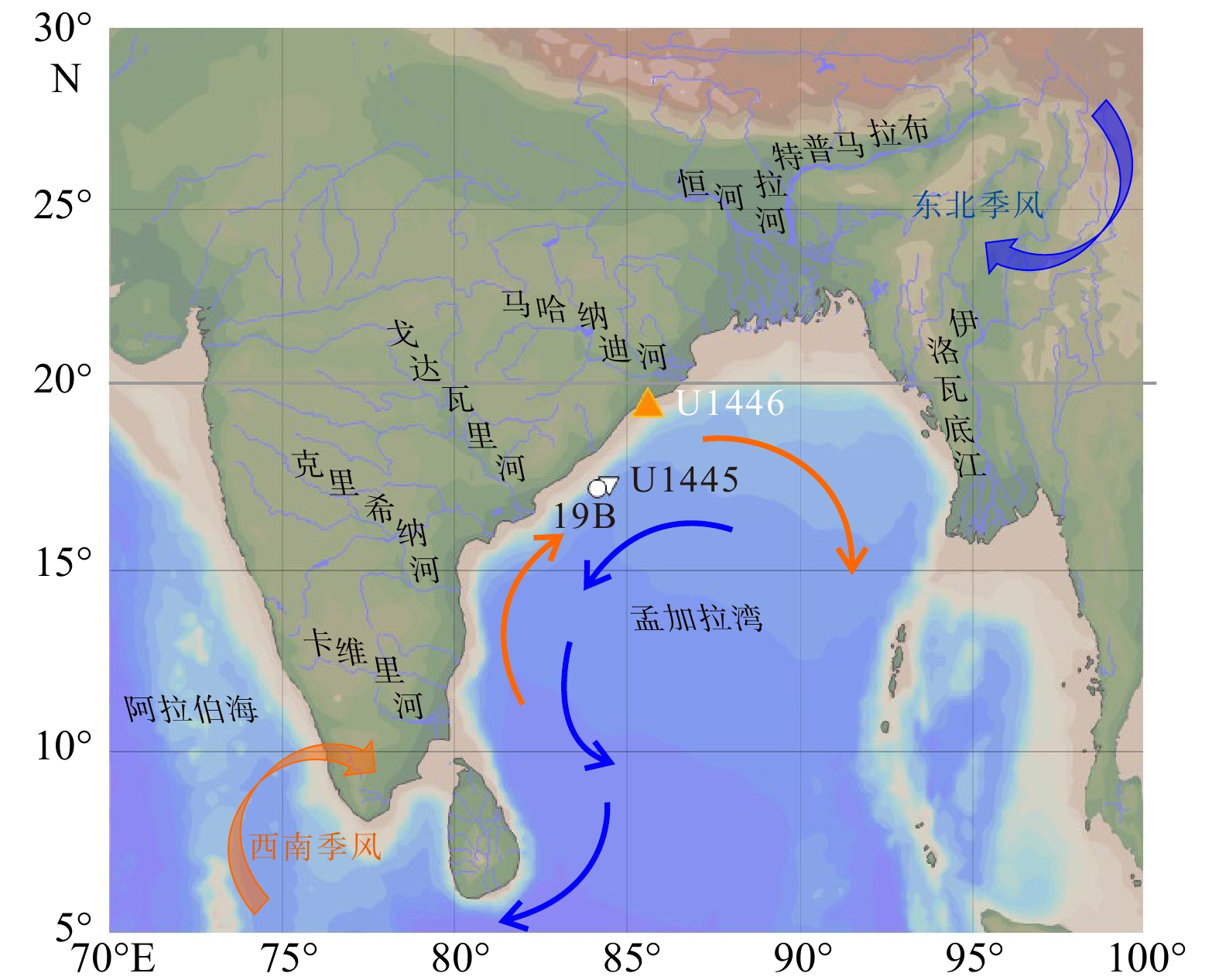
![]() 图 1 孟加拉湾的表层环流及周边主要河流橙色箭头代表西南夏季风环流,蓝色箭头代表东北冬季风环流。图形使用Ocean Data View绘制。Figure 1. Surface circulation in the Bay of Bengal and major rivers nearbyOrange arrows represent the southwest summer monsoon currents and blue arrows represent the northeast winter monsoon currents. The figure was created by Ocean Data View.
图 1 孟加拉湾的表层环流及周边主要河流橙色箭头代表西南夏季风环流,蓝色箭头代表东北冬季风环流。图形使用Ocean Data View绘制。Figure 1. Surface circulation in the Bay of Bengal and major rivers nearbyOrange arrows represent the southwest summer monsoon currents and blue arrows represent the northeast winter monsoon currents. The figure was created by Ocean Data View.1. 区域概况
孟加拉湾位于东北印度洋,是一个三面环陆的半封闭海湾,与印度、斯里兰卡、孟加拉国、缅甸和安达曼-尼科巴群岛接壤,是世界上最大的海湾,其面积达2.2×106 km2,占世界大洋面积的0.6%[14]。
孟加拉盆地东侧为安达曼-苏门答腊沟-弧-盆体系,西部与印度东部大陆边缘相邻,北靠孟加拉陆架,南部与中印度洋海盆和沃顿海盆相邻。在始新世之前,海盆物源主要来自西侧的印度半岛,始新世开始的喜马拉雅造山带逐渐占据优势,分别源自于喜马拉雅山脉南北坡的两条南亚大河——恒河(Ganga)和布拉马普特拉河(Brahmaputra)的陆源沉积物汇入孟加拉海盆,形成了世界上规模最大的深海扇复合体,即孟加拉和尼科巴深海扇系统。该深海扇从大陆架向海延伸至3000 km,最远端可至5°~7°S [15]。马哈纳迪盆地位于西北孟加拉湾19° 21′~23° 35′N、80° 30′~86° 50′E之间,面积约为1.42×105 km2。该沉积盆地从陆上延伸至海洋,形成于冈瓦纳古陆的晚侏罗纪分裂期间[16]。
孟加拉湾地处热带,具有明显的干湿两季,夏季盛行西南季风并且降雨充沛,冬季盛行东北季风(图1)。孟加拉湾表层海水温度总体较高且季节性变化较小,年平均温度为27~28.5 ℃[17]。在冬季,孟加拉湾表层海水温度呈现由北向南逐渐升高的趋势,北部最低表层海水温度约为25.5 ℃,南部约为29 ℃;夏季表层海水温度分布相对均一,约为28~29 ℃。
表层水的年平均盐度为30 ~34 psu不等,降水的季节性变化导致孟加拉湾表层水的盐度梯度随季节而发生变化。在夏季西南季风期间,由于降水增加,这种盐度梯度变得更加极端,可以从28 psu到36 psu[18]。恒河、布拉马普特拉河、克里希纳河(Krishna)、戈达瓦里河(Godavari)、马哈纳迪河(Mahanadi)、伊洛瓦底江(Irrawaddy)和卡维里河(Cauvery)等6条主要河流贡献了孟加拉湾淡水总量的60%,其中Ganga-Brahmaputra (G-B) 河系占44%[19]。每年河流水流量能达到2.95×1012 m3,并且由于降水量远超过蒸发量,导致孟加拉湾的上层海水由于盐度差异出现稳定的分层现象[8,19]。孟加拉湾表面的水平衡使得夏季季风期间净降水和径流增加63.7 cm,冬季季风期间净蒸发11.5 cm,导致年度净降水过剩[20]。
孟加拉湾北部的表层海水硅酸盐含量也表现出明显的季节性特点。夏季表层海水硅酸盐含量急剧上升,比冬季高出4~6倍以上。尤其是在G-B河系的入海口,硅酸盐浓度的变化极为明显(图2)。Miranda等[21]发现西北孟加拉湾硅酸盐的最大值和最小值分别出现在7月的夏季风期间和3月的冬季风期间。
![]() 图 2 孟加拉湾1995—2012年冬季、夏季表层海水温盐图以及硅酸盐浓度对比图图形使用Ocean Data View绘制,数据来源于World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de))。Figure 2. Salinity, temperature, and comparison of silicate concentration of surface sea water in the Bay of Bengal during winter and summer of 1955—2012Figures are created by Ocean Data View and data are from World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de)).
图 2 孟加拉湾1995—2012年冬季、夏季表层海水温盐图以及硅酸盐浓度对比图图形使用Ocean Data View绘制,数据来源于World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de))。Figure 2. Salinity, temperature, and comparison of silicate concentration of surface sea water in the Bay of Bengal during winter and summer of 1955—2012Figures are created by Ocean Data View and data are from World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de)).孟加拉湾的表层洋流主要由印度季风驱动,呈现出季节性方向反转的涡流形式[5]。东印度沿岸的孟加拉湾西部边界流在印度夏季风期间向北流动,在印度冬季风期间向南流动[22]。尽管沿印度半岛东部边缘出现了埃克曼(Ekman)驱动的沿岸上升流,但在夏季风期间,由于淡水大量注入导致的海水分层现象,使得上升流被限制在了离岸40 km以内[23]。
2. 材料与方法
2.1 研究材料
本次研究的材料为IODP353航次在印度季风的核心对流区孟加拉湾所获取的U1446站位岩心样品。U1446站位位于19°5′N、85°44′E,水深1430 m,离东印度海岸线大约75 km,在印度半岛东缘马哈纳迪盆地内。
U1446站位由U1446A、U1446B和U1446C 3个孔组成,3个孔的钻探深度分别为海底之下180.11、27.35和182.18 m。该站位岩心沉积物时代分布自更新世早期至全新世,为半远洋沉积,主要成分是岩屑以及少量的生物碎屑。沉积物主要由深灰色到浅灰色黏土所组成,含有不同比例的钙质超微化石、有孔虫以及生物硅。其中,岩屑成分平均占沉积物的70%,所占比例可从25%变化到85%。而生物成分主要是超微化石和有孔虫,含量变化从10%到25%不等。生物硅的含量一般较低,在特定的段落偶尔能观测到硅藻、放射虫和其他难以识别的生物硅质碎片[24]。在沉积物样品实际观察中,可发现钙质超微化石连续分布在U1446站位的沉积物中,而硅质微体化石却呈现零散的分布[24]。
本次研究选用A、C两孔的整合岩心,仅对其中71~142 m段进行研究,该段属于早更新世晚期至中更新世早期,采样间隔为45~50 cm,共采取了135个样品。
2.2 研究方法
样品的生物硅含量分析采取硅钼蓝分光光度计比色法[25],该方法误差小于3%。基本原理是先用H2O2和HCl溶液去除样品中的有机质和碳酸盐,再用Na2CO3碱性溶液将生物硅从沉积物中提取出来,然后与钼酸铵溶液反应生成硅钼黄,再加入抗坏血酸溶液还原为硅钼蓝。在一定浓度范围内,其蓝色与SiO2的浓度成正比,用722型分光光度计测得该溶液的吸光度,将吸光度带入回归方程计算得出生物硅的百分含量。生物硅含量分析的预处理与测试均在中国地质大学(北京)海洋学院完成。
为避免非硅质沉积稀释作用的影响,我们运用如下公式估算生物硅的沉积通量:
$$\begin{split}&\rm{\text {生物硅通量}}(g/cm^{2}\cdot ka)={\text {生物硅含量}}{(\text{%})}\times\\&\rm {\text {干密度}}(g/cm^{3})\times {\text {沉积速率}}(cm/ka)\end{split}$$ (1) $$\begin{split}&\rm {\text{干密度}}(g/cm^{3})={\text{湿密度}}(g/cm^{3})\times 1.540\;2-1.545\;3\\& (R^{2}=0.997\;2)\end{split}$$ (2) 通量公式(1)来自于Wang[26],干密度公式(2)来自于Phillips[9],U1446岩心样品湿密度数据通过伽马射线衰变获得,来源于IODP官网(OVERVIEW - Ocean Drilling Data (tamu.edu)),并通过插值法获得对应样品深度处的干密度值。
3. 年龄框架
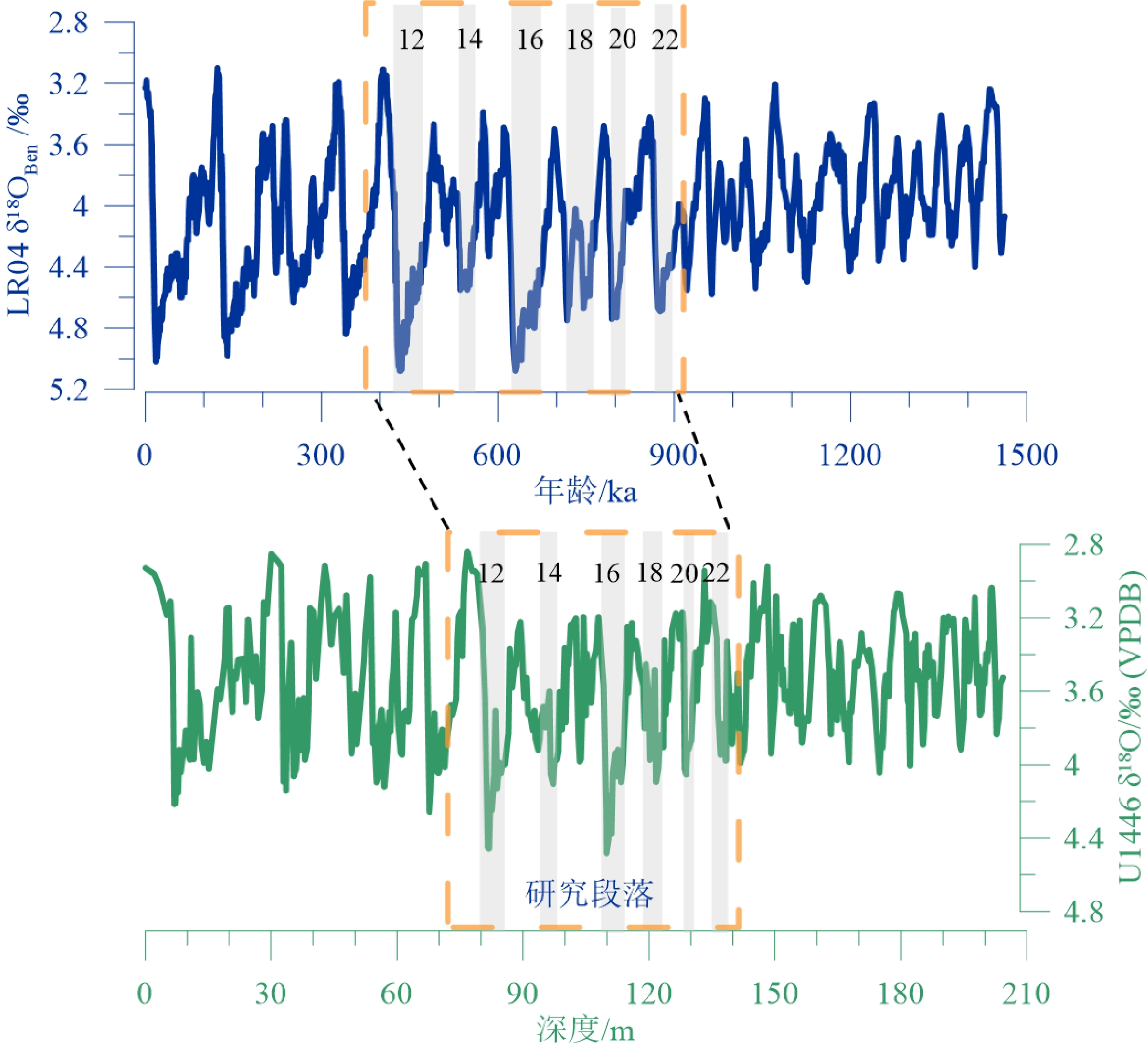
Clemens等[27]通过对U1446站位A、C两个孔整合岩心上部8 m的6个AMS14C年龄控制点,以及底栖有孔虫δ18O值与全球大洋LR04标准曲线进行对比,建立了U1446站位岩心的年代地层框架。
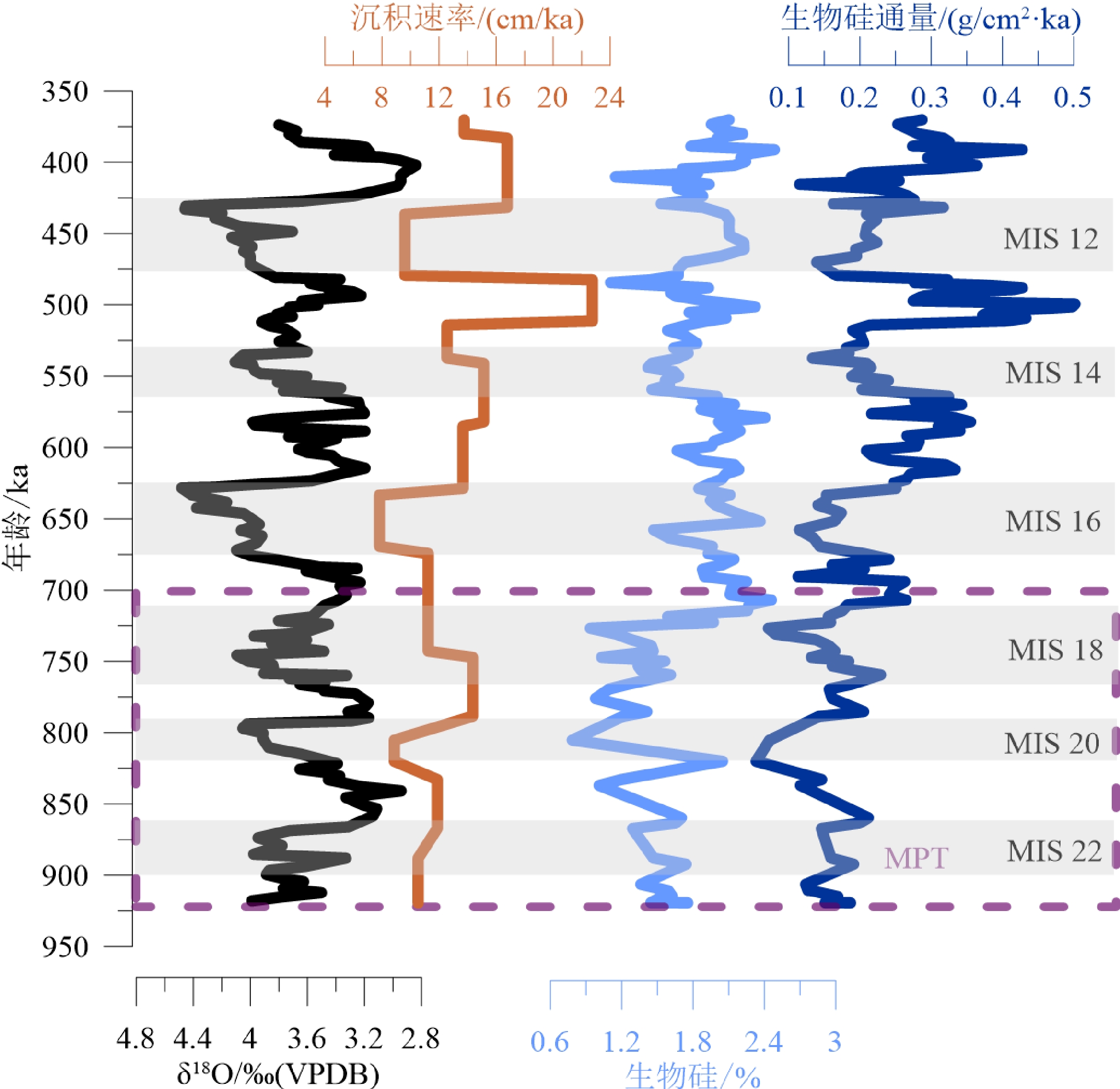
底栖有孔虫δ18O是从样品中大小为250~355 μm和大于355 μm(当250~355 μm的个体达不到分析要求的数量时)的底栖有孔虫中,挑选出Uvigerina spp.和Cibicidoides wuellerstorfi壳体,经超声清洗之后在MAT252稳定同位素质谱仪上测得[27]。所测结果与碳酸盐同位素标准NBS19进行校准,并以‰VPDB表示。U1446岩心的底栖有孔虫δ18O曲线如图3,其中方框指示本次研究的样品区间71~142 m,年龄为920~370 ka,包含氧同位素(Marine Isotope Stages, MIS)11—22期,样品的平均时间分辨率为4.1 ka。
![]()
4. 结果
西北孟加拉湾U1446站位920~370 ka期间生物硅含量总的变化范围为0.78%~2.48%,平均为1.79%,生物硅通量总的变化范围为0.05 ~0.50 g/cm2·ka,平均为0.23 g/cm2·ka(图4)。其中,MIS17期和MIS18期之间(约720 ka)是个明显的分界点,920~720 ka生物硅含量和通量整体相对较低且变化幅度小,平均值分别为1.40%和0.15 g/cm2·ka,生物硅通量在820 ka左右出现最低值,仅为0.05 g/cm2·ka;720~370 ka生物硅含量和通量迅速升高,平均值分别为1.91%和0.26 g/cm2·ka,且变化幅度加剧,生物硅通量在500 ka左右出现最高值,达0.50 g/cm2·ka。
5. 讨论
5.1 西北孟加拉湾表层硅质生物生产力
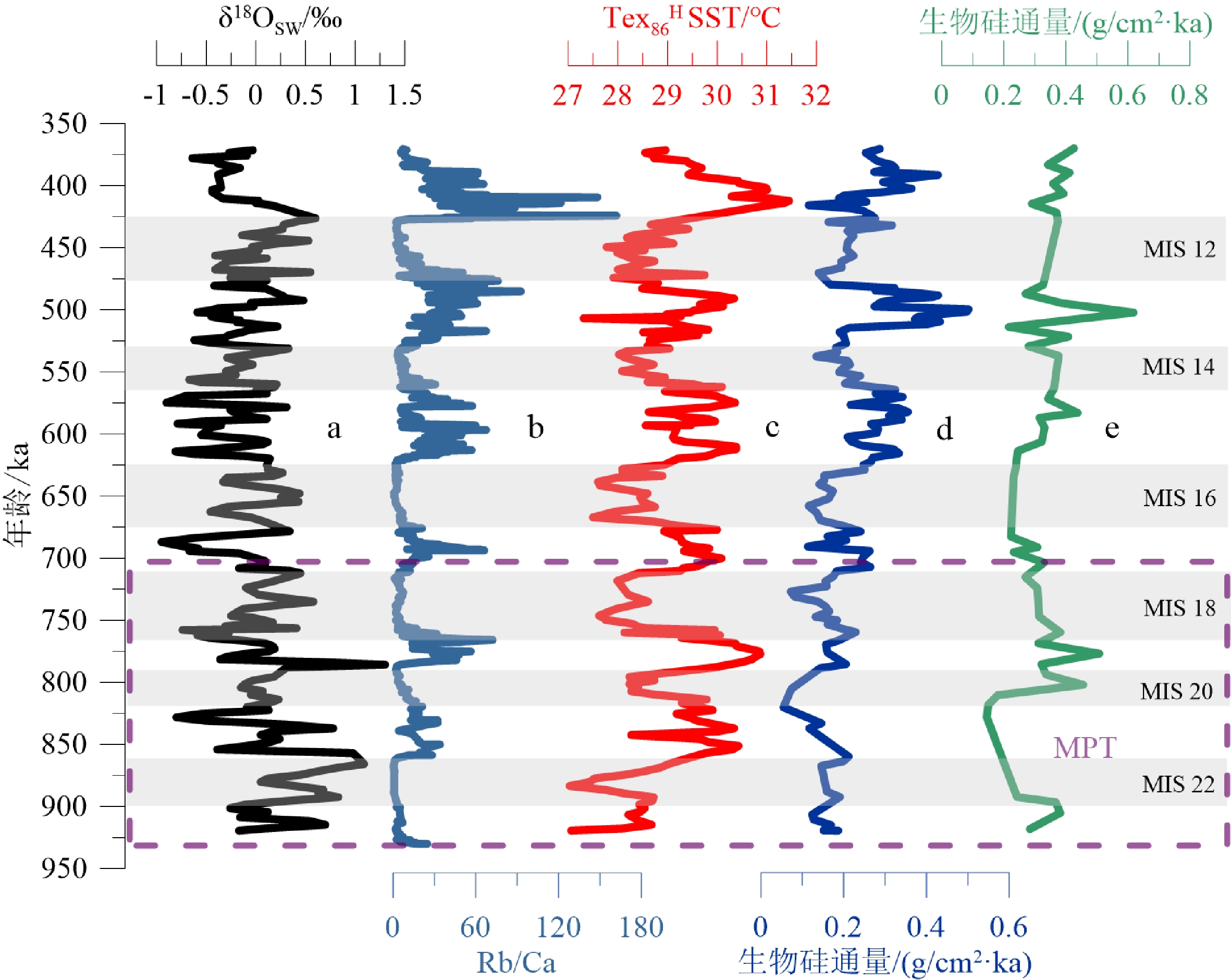
生源蛋白石又称生物硅,是由海洋硅质生物(如硅藻、放射虫等)死亡后的硅质骨骼降解形成的无定型蛋白石,因此沉积物中生物硅含量的高低可以用来反映海洋表层硅质生物的生产力[29]。海底沉积物中生物硅的含量主要受硅质骨架的供给、水体中硅质物质的溶解以及来自陆源输入物质的稀释作用影响。生物硅的沉积通量已排除了岩屑等陆源物质对沉积物中生物硅的稀释作用。Nelson等[4]认为生物硅在上层100 m的海水中至少溶解了50%,而在1000~3000 m水深范围则相当恒定,且硅质生物的溶解与钙质生物不同,不存在完全溶解的蛋白石补偿深度[30]。以前的研究[31]也证实,水体中只有很少比例的硅质骨骼发生溶解。前人的分析发现,海水中SiO2的溶解主要受温度和压力的影响,水温越高溶解度越大,压力越高溶解度越小[32]。U1446站位水深变化很小,因而压力的因素可以忽略,而该站位920~370 ka期间生物硅通量表现为间冰期时高冰期时低的特点,即水温越高生物硅通量越大(图5),指示溶解作用并没有对沉积物中生物硅通量的波动产生影响,推测该站位生物硅通量的变化主要受硅质生物生产力的控制。以前的研究也认为孟加拉湾地区的生物硅沉积通量是很好的表层海水硅质生产力指标 [11,33]。
![]() 图 5 季风指标与生物硅通量的比较a. U1446站位δ18Osw值,b. U1446站位Rb/Ca比值,c. Tex86H 换算得到U1446站位表层古水温变化,d. U1446站位生物硅通量,e. U1445站位生物硅通量。a-c数据来自于Clemens等[27]以及NOAA官网(https://www.ncei.noaa.gov/access/paleo-search/study/33213),e数据来源于Lee等[11]。Figure 5. Comparison of monsoon indicators to biogenic silica fluxesa: δ18Osw at Site U1446, b: Rb/Ca ratio at Site U1446, c: Variations in surface paleotemperature at Site U1446 from Tex86H converted data, d: biogenic silica fluxes (Data in a–c are from Clemens et al [27] and the NOAA official website (https://www. ncei.noaa.gov/access/paleo-search/study/33213)) , e: data are from Lee et al[11].
图 5 季风指标与生物硅通量的比较a. U1446站位δ18Osw值,b. U1446站位Rb/Ca比值,c. Tex86H 换算得到U1446站位表层古水温变化,d. U1446站位生物硅通量,e. U1445站位生物硅通量。a-c数据来自于Clemens等[27]以及NOAA官网(https://www.ncei.noaa.gov/access/paleo-search/study/33213),e数据来源于Lee等[11]。Figure 5. Comparison of monsoon indicators to biogenic silica fluxesa: δ18Osw at Site U1446, b: Rb/Ca ratio at Site U1446, c: Variations in surface paleotemperature at Site U1446 from Tex86H converted data, d: biogenic silica fluxes (Data in a–c are from Clemens et al [27] and the NOAA official website (https://www. ncei.noaa.gov/access/paleo-search/study/33213)) , e: data are from Lee et al[11].U1446站位生物硅沉积通量反映研究区在920~370 ka期间硅质生产力总体上呈现出逐渐上升的趋势(图4、5),其变化特征主要表现为:(1)在长时间尺度上,MPT(920~700 ka)期间生物硅通量较低,在820 ka左右出现最小值,变化幅度也偏弱,指示硅质生产力较低且波动不是很强烈。在MPT之后,生物硅通量与沉积速率出现明显上升的趋势,特别是在630 ka之后,硅质生产力明显增强且波动明显。相邻U1445站位较U1446站位的研究时间分辨率较低,但同一阶段的生物硅通量[11]也表现出类似的变化趋势,且该站位生物硅通量和有机碳通量变化趋势一致[11]。(2)在轨道时间尺度上,U1446站位生物硅通量呈现出明显的冰期间冰期旋回性:冰期时生物硅通量下降,指示表层硅质生产力较低;而间冰期生物硅通量上升,指示表层硅质生产力上升。且在MPT阶段间冰期MIS17、MIS19、MIS21和冰期MIS18、MIS20、MIS22之间的古生产力差别较小,而在MPT之后,间冰期MIS11、MIS13、MIS15和冰期MIS12、MIS14、MIS16之间的古生产力差别迅速加大。位于西北孟加拉湾的19B站位80 ka以来的硅质生产力变化[12],虽然和本次研究不是同一个时间段,但也表现出同样的冰期-间冰期旋回性。
![]() 图 4 U1446站位920~370 ka沉积速率、生物硅含量及通量与δ18O/‰(VPDB)的对比Figure 4. Comparison in sedimentation rate, biogenic silica content, fluxes, and δ18O/‰ (VPDB) of Site U1446, 920~370 ka
图 4 U1446站位920~370 ka沉积速率、生物硅含量及通量与δ18O/‰(VPDB)的对比Figure 4. Comparison in sedimentation rate, biogenic silica content, fluxes, and δ18O/‰ (VPDB) of Site U1446, 920~370 ka5.2 西北孟加拉湾硅质生产力对印度季风活动的响应
西北孟加拉湾是印度季风降水的核心区域,其表层海水盐度、温跃层深度、营养物质供应、表层水生产力的变化均与印度季风活动密切相关。印度夏季风增强时,不仅海洋表面接受强烈的季风降水,大量的降水也在陆地河流集水并由径流带入西北孟加拉湾,导致研究区表层海水盐度下降。为讨论U1446站位的古海水盐度变化,本次研究使用去除了冰体积效应和表层水温度效应的表层海水氧同位素指标(δ18Osw),它与表层海水盐度呈线性相关[33-34],δ18Osw高值时指示表层水盐度上升,低值时指示表层水盐度下降,是孟加拉湾地区夏季风降水的重要指标[27]。
Rb(铷)是长英石中常见的碱性金属元素,它会随着径流量的提升、细粒岩屑物质输入量的增加而增加[27]。以前的研究发现,西北孟加拉湾碳酸钙的沉积通量在印度夏季风增强时下降,减弱时上升[9,26],Phillips等[9]认为是夏季风增强引起的强降水使得孟加拉湾上层海水出现严重的分层现象,以致于海水垂直交换受阻,上升流活动受到抑制,底层水的营养物质难以输送到表层,导致钙质生物生产力下降。因此,孟加拉湾地区沉积物中的Rb/Ca比值随夏季风增强淡水径流量增加而升高,夏季风减弱淡水径流减少而下降,也被作为印度夏季风活动的替代性指标[27]。
U1446站位δ18Osw值和Rb/Ca比值[27]显示(图5),在MPT(920~700 ka)阶段,δ18Osw值较重而Rb/Ca比值则较低,指示研究区表层水盐度较高、陆表径流量不大,表明该阶段研究区印度夏季风活动较弱,而生物硅通量显示的硅质生物生产力较低。而在MPT之后的700~370 ka阶段,δ18Osw值减轻,反映表层海水盐度有下降的趋势,且Rb/Ca比值明显升高,指示陆表径流量上升,显示研究区印度夏季风活动增强,而生物硅通量则显示硅质生物生产力明显上升。另外,U1446站位δ18Osw值和Rb/Ca比值[27]都显示出明显的冰期间冰期旋回性,与生物硅通量变化趋势一致,指示间冰期夏季风增强、降水和径流量增加时硅质生产力上升,冰期夏季风减弱、降水和径流量下降时硅质生产力下降;且在MPT之后,硅质生产力变化的幅度显著加强,且整体高于MPT时期的平均值。
虽然太阳辐射量、温度、营养物质、海水分层现象等都是造成大洋生产力高低变化的因素,但其中最关键的控制因素还是营养物质的丰富程度。以前的研究指出,印度洋钙质生物生产力受营养元素N的输入量控制,当印度夏季风增强时,大量的淡水输入导致海水分层,使得混合层深度浅于营养跃层顶部,限制了下部富营养元素N的海水混合和上升进入透光带,因而研究区钙质生产力下降[9,11,33]。然而,硅质生产力则更多地受陆地径流带来的营养元素Si和P的控制[33-34],透光带低的N/Si比值更有利于硅藻的发育[35]。
由于孟加拉湾周边大量的河流汇入,河流带来的淡水占据孟加拉湾总淡水量的60%[19]。在夏季,印度夏季风所带来的直接降水以及河流汇入水量占据研究区全年淡水量的90 %[36]。而G-B河系给孟加拉湾的营养盐供应和全球河系的对比都非常显著。每年通过G-B河系供应给大洋的二氧化硅占全球河流输入海洋的4%以上[37-38]。无机磷输入的平均浓度高于全球河流的平均值,输入到孟加拉湾的量占据全球河流年输出的10 %[39]。汇入孟加拉湾的巨型河流会提供大量的溶解硅酸盐[40],硅酸盐又是硅藻生长繁殖的决定性因素[10]。因此,径流输入的营养盐对于孟加拉湾表层海水浮游生物的生长繁殖起到重要的作用。印度夏季风的增强会导致地表径流显著升高,伴随而来的大量营养盐使得海水表层硅质生物生产力升高。因此西北孟加拉湾地区常常出现碳酸钙通量反映的钙质生物生产力和硅质生物生产力变化呈现反相关关系[11-12]的特点。
5.3 生物硅变化对轨道周期的响应
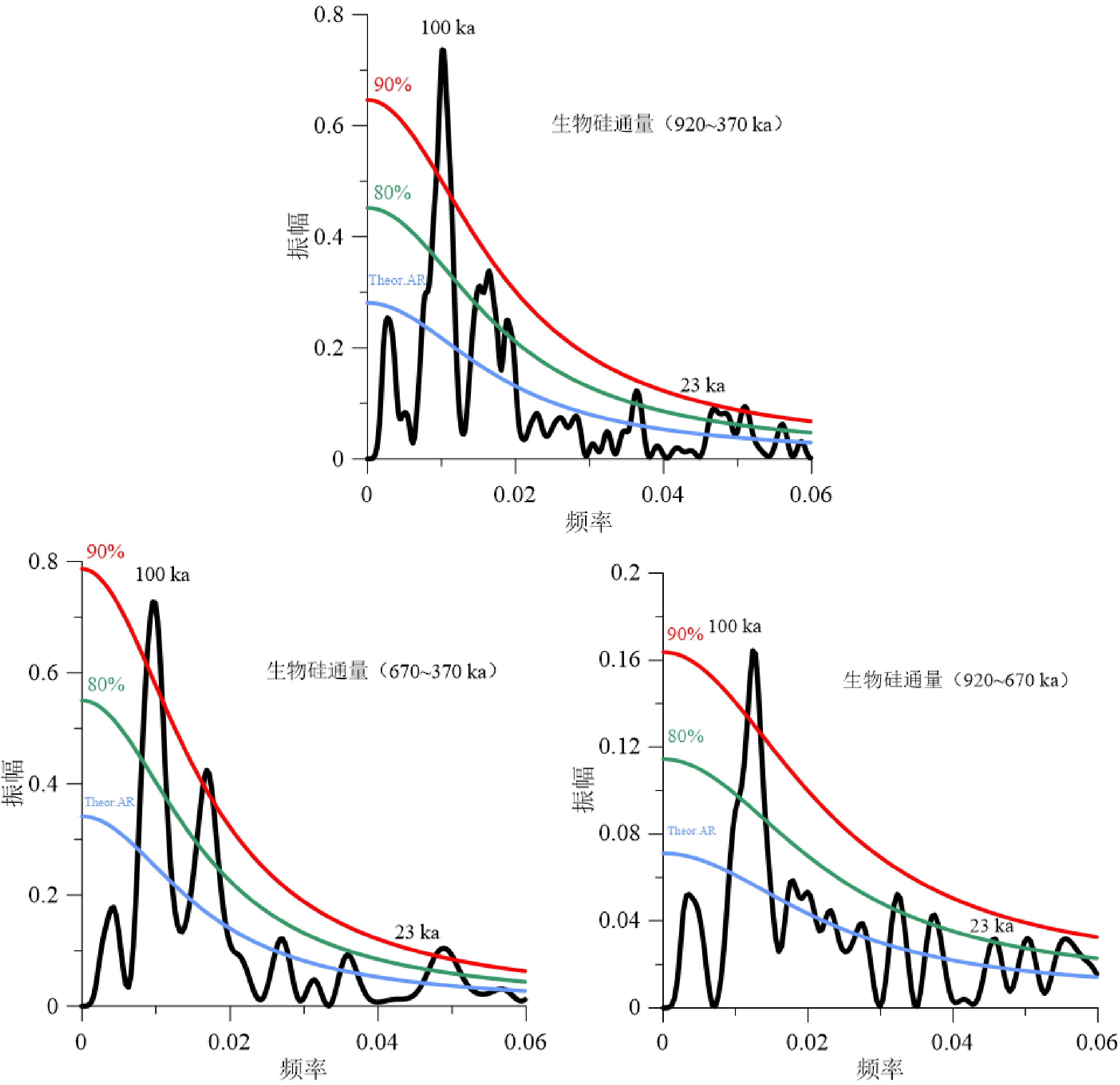
通过对U1446站位920~370 ka整个阶段以及MPT和MPT之后分阶段的生物硅通量的频谱分析结果表明,研究区生物硅通量整体上以100 ka偏心率周期占主导地位,41 ka的斜率周期不明显,与降水以及径流指标(δ18Osw,Rb/Ca)的频谱结果[27]比较一致。值得注意的是,δ18Osw有明显的岁差周期,反映夏季风降水的热带驱动,生物硅通量23 ka的岁差周期也较为明显(图6)。在MPT(920~670 ka)阶段,生物硅通量以偏心率周期为主导,斜率周期和岁差周期虽然置信度不高,但也都有出现。MPT之后(670~370 ka),仍以偏心率周期为主导,岁差周期也很明显,二者置信度在90 %以上,但41 ka斜率周期不明显。
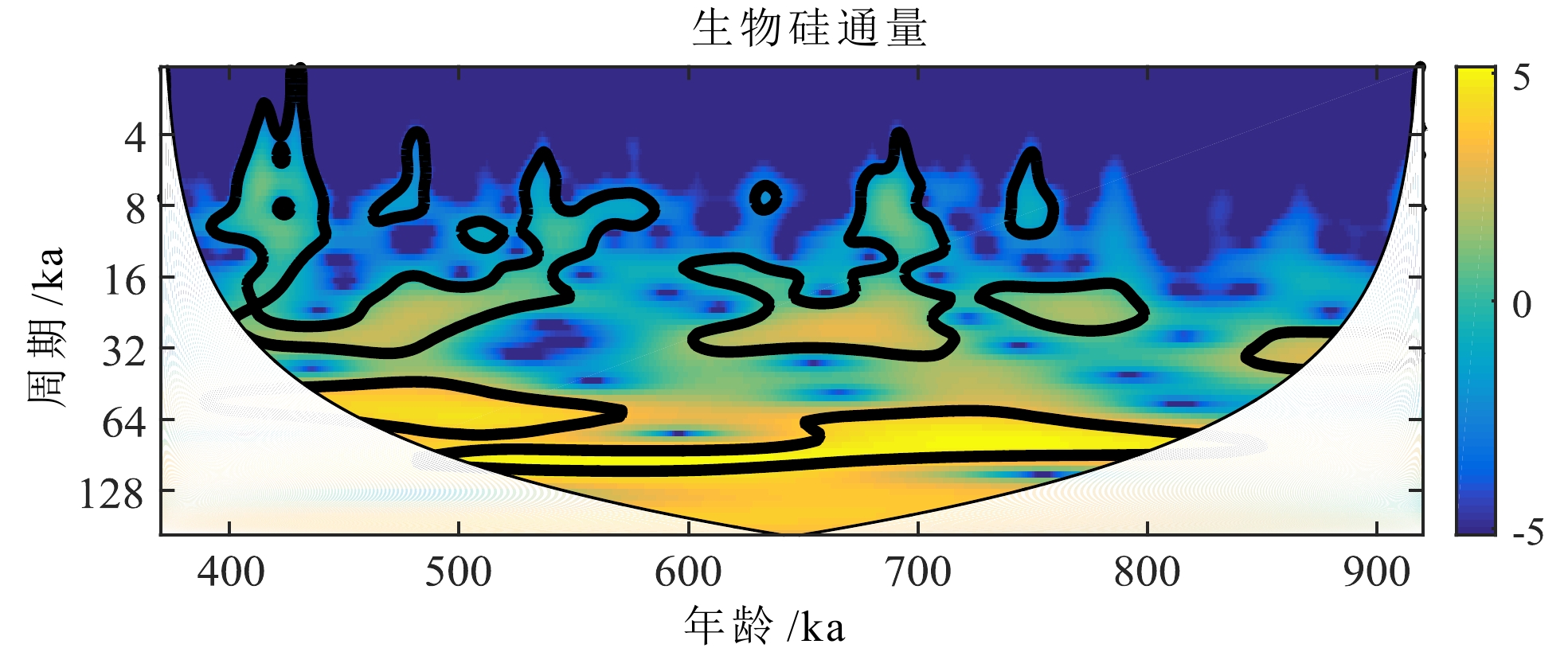
对U1446站位920~370 ka期间连续小波分析的结果显示(图7),生物硅通量变化对轨道周期的响应非常明显,表现为偏心率周期连续,在650~500 ka之间较强,650 ka之前略有偏离;岁差周期贯穿始终,但不连续;斜率周期全程较弱,主要出现在920~850 ka,即MPT期间。
![]() 图 7 孟加拉湾U1446站位920~370 ka生物硅通量小波分析Figure 7. Wavelet analysis of biogenic silica fluxes at Site U1446, Bay of Bengal, 920~370 ka
图 7 孟加拉湾U1446站位920~370 ka生物硅通量小波分析Figure 7. Wavelet analysis of biogenic silica fluxes at Site U1446, Bay of Bengal, 920~370 ka不同的轨道参数在不同的纬度作用不同。Short等[41]发现低纬区主要响应23 ka的岁差周期,而偏心率则是通过对岁差的调幅作用进入气候系统,因此低纬地区受岁差和偏心率的综合驱动更大。
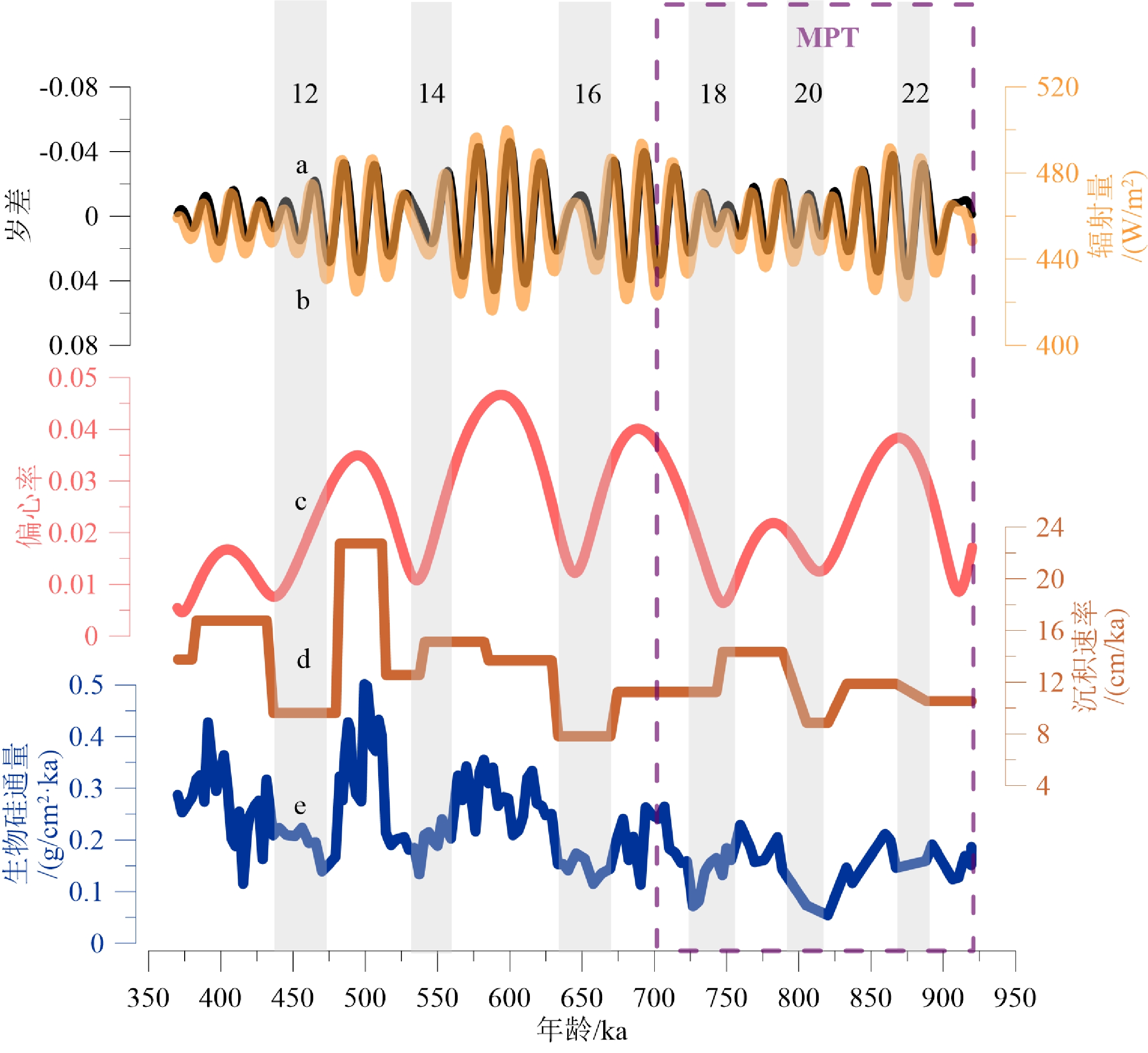
通过对比U1446站位920~370 ka期间生物硅通量和地球轨道参数、北纬15°夏季太阳辐射量的变化(图8)发现,当偏心率增大时,岁差变幅和太阳辐射量也较大,这样会导致地球气候效应即季节差异随之加大,表现为全球季风的变幅和强度的增加[42],尤其是在偏心率、岁差和太阳辐射量的周期同时出现高峰时,此时的生物硅通量呈上升趋势且幅度剧烈。因而U1446站位出现冰期-间冰期硅质生产力的明显差异,且生物硅通量的小波分析图中也明显可见650~500 ka之间更清晰的偏心率周期(图7)。另外,U1446站位生物硅通量出现更明显的岁差周期,可能与藻类的光合作用也受到太阳辐射量的影响有关。
![]() 图 8 生物硅含量及通量和地球轨道参数的比较a. 岁差,b. 北纬15°夏季(7月)月均太阳辐射,c. 偏心率,d. 沉积速率,e. 生物硅通量。a-c数据来源于Berger[47],d-e数据均来自U1446孔。Figure 8. Comparison of biogenic silica content and fluxes and the Earth orbital parametersa: precession, b: average monthly solar radiation in summer (July) at 15°N, c: eccentricity, d: sedimentation rate, e: biogenic silica fluxes. Data in a–c are derived from Berger [47], d–e data are from Site U1446.
图 8 生物硅含量及通量和地球轨道参数的比较a. 岁差,b. 北纬15°夏季(7月)月均太阳辐射,c. 偏心率,d. 沉积速率,e. 生物硅通量。a-c数据来源于Berger[47],d-e数据均来自U1446孔。Figure 8. Comparison of biogenic silica content and fluxes and the Earth orbital parametersa: precession, b: average monthly solar radiation in summer (July) at 15°N, c: eccentricity, d: sedimentation rate, e: biogenic silica fluxes. Data in a–c are derived from Berger [47], d–e data are from Site U1446.大量的研究发现降水与偏心率周期及太阳辐射量密切相关,且对冰盖的减少、温室气体的增多敏感[43-44],西北孟加拉湾降水量和径流量的最大值都发生在北半球夏季太阳辐射量的最大值,以偏心率周期为主导[27],降水和径流量的提升使得汇入西北孟加拉湾的硅酸盐含量增加,硅藻大量繁殖,生物硅含量及通量上升。
前人对冰盖的扩张和收缩对沉积物中生物硅的影响已做过很多研究[45-46],通过对比热带地区(孟加拉湾)与亚寒带地区(白令海)的生物硅变化发现[11,44,46],两区域的生物硅通量都在MPT时期有显著下降,虽然表面上看影响因素不同,例如亚寒带生物硅在MPT时期下降的原因是由于冰盖扩张所导致的,而西北孟加拉湾则主要受季风降水和径流量的影响,但它们是否都与更新世地球轨道参数变化导致的全球冰体积和温室气体有关,其动力机制还有待进一步研究。
6. 结论
(1)IODP353航次U1446站位生物硅含量及通量的变化结果显示,西北孟加拉湾在920~370 ka期间硅质生产力总体上呈现出逐渐上升的趋势。在长时间尺度上,MPT期间硅质生产力较低,MPT之后硅质生产力明显上升且波动强烈;在轨道尺度上,硅质生产力则呈现出明显的冰期-间冰期旋回性,表现为冰期时下降,间冰期时上升。
(2)对比该站位印度夏季风降水和径流指标发现,研究区夏季风增强时硅质生产力上升,减弱时硅质生产力下降,指示该区硅质生产力更多地受陆地径流带来的营养元素Si和P的控制。夏季风增强时导致孟加拉湾降水以及周边河流径流量剧增,河流提供的营养物质使得研究区硅质生产力迅速升高,夏季风减弱时营养盐的减少使得硅质生产力下降。
(3)U1446站位生物硅通量的频谱和小波分析结果显示,生物硅通量不同程度上受到了地球轨道周期的控制,尤其是在MPT之后受到偏心率周期的调控较为明显。
致谢:国际大洋钻探IODP 353航次的全体科学家和工作人员的辛苦付出,中国IODP办公室提供航次资助,同济大学王汝建教授指导生物硅实验,在此一并致谢!
-
![]()
图 1 孟加拉湾的表层环流及周边主要河流
橙色箭头代表西南夏季风环流,蓝色箭头代表东北冬季风环流。图形使用Ocean Data View绘制。
Figure 1. Surface circulation in the Bay of Bengal and major rivers nearby
Orange arrows represent the southwest summer monsoon currents and blue arrows represent the northeast winter monsoon currents. The figure was created by Ocean Data View.
![]()
图 2 孟加拉湾1995—2012年冬季、夏季表层海水温盐图以及硅酸盐浓度对比图
图形使用Ocean Data View绘制,数据来源于World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de))。
Figure 2. Salinity, temperature, and comparison of silicate concentration of surface sea water in the Bay of Bengal during winter and summer of 1955—2012
Figures are created by Ocean Data View and data are from World Ocean Atlas (ODV: World Ocean Atlas 2013 (awi.de)).
![]()
![]()
图 5 季风指标与生物硅通量的比较
a. U1446站位δ18Osw值,b. U1446站位Rb/Ca比值,c. Tex86H 换算得到U1446站位表层古水温变化,d. U1446站位生物硅通量,e. U1445站位生物硅通量。a-c数据来自于Clemens等[27]以及NOAA官网(https://www.ncei.noaa.gov/access/paleo-search/study/33213),e数据来源于Lee等[11]。
Figure 5. Comparison of monsoon indicators to biogenic silica fluxes
a: δ18Osw at Site U1446, b: Rb/Ca ratio at Site U1446, c: Variations in surface paleotemperature at Site U1446 from Tex86H converted data, d: biogenic silica fluxes (Data in a–c are from Clemens et al [27] and the NOAA official website (https://www. ncei.noaa.gov/access/paleo-search/study/33213)) , e: data are from Lee et al[11].
![]()
图 4 U1446站位920~370 ka沉积速率、生物硅含量及通量与δ18O/‰(VPDB)的对比
Figure 4. Comparison in sedimentation rate, biogenic silica content, fluxes, and δ18O/‰ (VPDB) of Site U1446, 920~370 ka
![]()
图 7 孟加拉湾U1446站位920~370 ka生物硅通量小波分析
Figure 7. Wavelet analysis of biogenic silica fluxes at Site U1446, Bay of Bengal, 920~370 ka
![]()
图 8 生物硅含量及通量和地球轨道参数的比较
a. 岁差,b. 北纬15°夏季(7月)月均太阳辐射,c. 偏心率,d. 沉积速率,e. 生物硅通量。a-c数据来源于Berger[47],d-e数据均来自U1446孔。
Figure 8. Comparison of biogenic silica content and fluxes and the Earth orbital parameters
a: precession, b: average monthly solar radiation in summer (July) at 15°N, c: eccentricity, d: sedimentation rate, e: biogenic silica fluxes. Data in a–c are derived from Berger [47], d–e data are from Site U1446.
-
[1] 汪品先, 翦知湣, 赵泉鸿, 等. 南海演变与季风历史的深海证据[J]. 科学通报, 2003, 48(23):2549-2561 doi: 10.1360/03wd0156 WANG Pinxian, JIAN Zhimin, ZHAO Quanhong, et al. Evolution of the South China Sea and monsoon history revealed in deep-sea records [J]. Chinese Science Bulletin, 2003, 48(23): 2549-2561. doi: 10.1360/03wd0156
[2] Felix M. A comparison of equations commonly used to calculate organic carbon content and marine palaeoproductivity from sediment data [J]. Marine Geology, 2014, 347: 1-11. doi: 10.1016/j.margeo.2013.10.006
[3] Xu Y H, Wang L, Lai Z K, et al. The biogenic silica variation and paleoproductivity evolution in the eastern Indian Ocean during the past 20 0000 a [J]. Acta Oceanologica Sinica, 2019, 38(1): 78-84. doi: 10.1007/s13131-019-1372-z
[4] Nelson D M, Tréguer P, Brzezinski M A, et al. Production and dissolution of biogenic silica in the ocean: revised global estimates, comparison with regional data and relationship to biogenic sedimentation [J]. Global Biogeochemical Cycles, 1995, 9(3): 359-372. doi: 10.1029/95GB01070
[5] Madhupratap M, Gauns M, Ramaiah N, et al. Biogeochemistry of the Bay of Bengal: physical, chemical and primary productivity characteristics of the central and western Bay of Bengal during summer monsoon 2001 [J]. Deep Sea Research Part II:Topical Studies in Oceanography, 2003, 50(5): 881-896. doi: 10.1016/S0967-0645(02)00611-2
[6] Cortese G, Gersonde R, Hillenbrand C D, et al. Opal sedimentation shifts in the World Ocean over the last 15 Myr [J]. Earth and Planetary Science Letters, 2004, 224(3-4): 509-527. doi: 10.1016/j.jpgl.2004.05.035
[7] Volk T, Hoffert M I. Ocean carbon pumps: analysis of relative strengths and efficiencies in ocean-driven atmospheric CO2 changes[M]//Sundquist E T, Broecker W S. The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present. Washington: American Geophysical Union, 1985: 99-110.
[8] Prasanna Kumar S, Nuncio M, Ramaiah N, et al. Eddy-mediated biological productivity in the Bay of Bengal during fall and spring intermonsoons [J]. Deep Sea Research Part I:Oceanographic Research Papers, 2007, 54(9): 1619-1640. doi: 10.1016/j.dsr.2007.06.002
[9] Phillips S C, Johnson J E, Giosan L, et al. Monsoon-influenced variation in productivity and lithogenic sediment flux since 110 ka in the offshore Mahanadi Basin, northern Bay of Bengal [J]. Marine and Petroleum Geology, 2014, 58: 502-525. doi: 10.1016/j.marpetgeo.2014.05.007
[10] Prasanna Kumar S, Muraleedharan P M, Prasad T G, et al. Why is the Bay of Bengal less productive during summer monsoon compared to the Arabian Sea? [J]. Geophysical Research Letters, 2002, 29(24): 2235.
[11] Lee J, Kim S, Khim B K. A paleoproductivity shift in the northwestern Bay of Bengal (IODP Site U1445) across the Mid-Pleistocene transition in response to weakening of the Indian summer monsoon [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 560: 110018. doi: 10.1016/j.palaeo.2020.110018
[12] Ota Y, Kuroda J, Yamaguchi A, et al. Monsoon-influenced variations in plankton community structure and upper-water column stratification in the western Bay of Bengal during the past 80 ky [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 521: 138-150. doi: 10.1016/j.palaeo.2019.02.020
[13] Clark P U, Archer D, Pollard D, et al. The middle Pleistocene transition: characteristics, mechanisms, and implications for long-term changes in atmospheric pCO2 [J]. Quaternary Science Reviews, 2006, 25(23-24): 3150-3184. doi: 10.1016/j.quascirev.2006.07.008
[14] LaFond E C. Bay of Bengal[M]//Fairbridge R W. The Encyclopedia of Oceanography. New York: Van Nostrand Reinhold Co. , 1966: 110-118.
[15] Curray J R, Emmel F J, Moore D G, et al. Structure, tectonics, and geological history of the northeastern Indian ocean[M]//Nairn A E M, Stehli F G. The Ocean Basins and Margins: The Indian Ocean. New York: Springer, 1982: 399-450.
[16] Subrahmanyam V, Subrahmanyam A S, Murty G P S, et al. Morphology and tectonics of Mahanadi Basin, northeastern continental margin of India from geophysical studies [J]. Marine Geology, 2008, 253(1-2): 63-72. doi: 10.1016/j.margeo.2008.04.007
[17] Chatterjee A, Shankar D, Shenoi S S C, et al. A new atlas of temperature and salinity for the North Indian Ocean [J]. Journal of Earth System Science, 2012, 121(3): 559-593. doi: 10.1007/s12040-012-0191-9
[18] Boyer T P, Antonov J I, Baranova O K, et al. World ocean database 2013[R]. Silver Spring: National Oceanographic Data Center Ocean Climate Laboratory, 2013.
[19] Da Silva R, Mazumdar A, Mapder T, et al. Salinity stratification controlled productivity variation over 300 ky in the Bay of Bengal [J]. Scientific Reports, 2017, 7(1): 14439. doi: 10.1038/s41598-017-14781-3
[20] Varkey M J, Murty V S N, Suryanarayana A. Physical oceanography of the bay of Bengal and Andaman sea [J]. Oceanography and Marine Biology:An Annual Review, 1996, 34: 1-70.
[21] Miranda J, Baliarsingh S K, Lotliker A A, et al. Long-term trend and environmental determinants of phytoplankton biomass in coastal waters of northwestern Bay of Bengal [J]. Environmental Monitoring and Assessment, 2020, 192(1): 55. doi: 10.1007/s10661-019-8033-8
[22] Shankar D, McCreary J P, Han W, et al. Dynamics of the East India Coastal Current: 1. Analytic solutions forced by interior Ekman pumping and local alongshore winds [J]. Journal of Geophysical Research:Oceans, 1996, 101(C6): 13975-13991. doi: 10.1029/96JC00559
[23] Shetye S R, Shenoi S S C, Gouveia A D, et al. Wind-driven coastal upwelling along the western boundary of the Bay of Bengal during the southwest monsoon [J]. Continental Shelf Research, 1991, 11(11): 1397-1408. doi: 10.1016/0278-4343(91)90042-5
[24] Clemens S C, Kuhnt W, LeVay L J, et al. Indian monsoon rainfall[C]//Proceedings of the International Ocean Discovery Program. College Station, TX: International Ocean Discovery Program, 2016: 1-15.
[25] Mortlock R A, Froelich P N. A simple method for the rapid determination of biogenic opal in pelagic marine sediments [J]. Deep Sea Research Part A. Oceanographic Research Papers, 1989, 36(9): 1415-1426. doi: 10.1016/0198-0149(89)90092-7
[26] Wang D, Ding X, Bassinot F. Observations of contrasted glacial-interglacial dissolution of foraminifera above the lysocline in the Bay of Bengal, northeastern Indian Ocean [J]. Acta Oceanologica Sinica, 2021, 40(1): 155-161. doi: 10.1007/s13131-021-1821-3
[27] Clemens S C, Yamamoto M, Thirumalai K, et al. Remote and local drivers of Pleistocene South Asian summer monsoon precipitation: a test for future predictions [J]. Science Advances, 2021, 7(23): eabg3848. doi: 10.1126/sciadv.abg3848
[28] Lisiecki L E, Raymo M E. A pliocene-pleistocene stack of 57 globally distributed benthic δ18O records [J]. Paleoceanography, 2005, 20(1): PA1003.
[29] 王汝建, 肖文申, 向霏, 等. 北冰洋西部表层沉积物中生源组分及其古海洋学意义[J]. 海洋地质与第四纪地质, 2007, 27(6):61-69 WANG Rujian, XIAO Wenshen, XIANG Fei, et al. Distribution pattern of biogenic components in surface sediments of the western arctic ocean and their paleoceanographic implications [J]. Marine Geology & Quaternary Geology, 2007, 27(6): 61-69.
[30] Berger W H. Biogenous deep sea sediments: production, preservation and interpretation[M]//Riley J P, Chester R. Treatise on Chemical Oceanography 5. San Diego, California: Academic, 1976: 265-388.
[31] Ragueneau O, Tréguer P, Leynaert A, et al. A review of the Si cycle in the modern ocean: recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy [J]. Global and Planetary Change, 2000, 26(4): 317-365. doi: 10.1016/S0921-8181(00)00052-7
[32] 同济大学海洋地质系. 古海洋学概论[M]. 上海: 同济大学出版社, 1989: 1-316 Department of Marine Geology, Tongji University. Introduction to Paleoceanography[M]. Shanghai: Tongji University Press, 1989: 1-316.
[33] Liu S F, Zhang H, Cao P, et al. Paleoproductivity evolution in the northeastern Indian Ocean since the last glacial maximum: evidence from biogenic silica variations [J]. Deep Sea Research Part I:Oceanographic Research Papers, 2021, 175: 103591. doi: 10.1016/j.dsr.2021.103591
[34] Achyuthan H, Deshpande R D, Rao M S, et al. Stable isotopes and salinity in the surface waters of the Bay of Bengal: implications for water dynamics and palaeoclimate [J]. Marine Chemistry, 2013, 149: 51-62. doi: 10.1016/j.marchem.2012.12.006
[35] Paul J T, Ramaiah N, Sardessai S. Nutrient regimes and their effect on distribution of phytoplankton in the Bay of Bengal [J]. Marine Environmental Research, 2008, 66(3): 337-344. doi: 10.1016/j.marenvres.2008.05.007
[36] Chakrapani G J, Subramanian V. Factors controlling sediment discharge in the Mahanadi River basin, India [J]. Journal of Hydrology, 1990, 117(1-4): 169-185. doi: 10.1016/0022-1694(90)90091-B
[37] Georg R B, West A J, Basu A R, et al. Silicon fluxes and isotope composition of direct groundwater discharge into the Bay of Bengal and the effect on the global ocean silicon isotope budget [J]. Earth and Planetary Science Letters, 2009, 283(1-4): 67-74. doi: 10.1016/j.jpgl.2009.03.041
[38] Laruelle G G, Roubeix V, Sferratore A, et al. Anthropogenic perturbations of the silicon cycle at the global scale: key role of the land–ocean transition [J]. Global Biogeochemical Cycles, 2009, 23(4): GB4031.
[39] Datta D K, Gupta L P, Subramanian V. Distribution of C, N and P in the sediments of the Ganges-Brahmaputra-Meghna river system in the Bengal basin [J]. Organic Geochemistry, 1999, 30(1): 75-82. doi: 10.1016/S0146-6380(98)00203-4
[40] Sarma V V S S, Rao G D, Viswanadham R, et al. Effects of freshwater stratification on nutrients, dissolved oxygen, and phytoplankton in the Bay of Bengal [J]. Oceanography, 2016, 29(2): 222-231. doi: 10.5670/oceanog.2016.54
[41] Short D A, Mengel J G, Crowley T J, et al. Filtering of milankovitch cycles by earth's geography [J]. Quaternary Research, 1991, 35(2): 157-173. doi: 10.1016/0033-5894(91)90064-C
[42] Wang P X, Tian J, Lourens L J. Obscuring of long eccentricity cyclicity in Pleistocene oceanic carbon isotope records [J]. Earth and Planetary Science Letters, 2010, 290(3-4): 319-330. doi: 10.1016/j.jpgl.2009.12.028
[43] Araya-Melo P A, Crucifix M, Bounceur N. Global sensitivity analysis of the Indian monsoon during the Pleistocene [J]. Climate of the Past, 2015, 11(1): 45-61. doi: 10.5194/cp-11-45-2015
[44] Erb M P, Jackson C S, Broccoli A J. Using single-forcing GCM simulations to reconstruct and interpret Quaternary climate change [J]. Journal of Climate, 2015, 28(24): 9746-9767. doi: 10.1175/JCLI-D-15-0329.1
[45] Kim S, Takahashi K, Khim B K, et al. Biogenic opal production changes during the Mid-Pleistocene Transition in the Bering Sea (IODP Expedition 323 Site U1343) [J]. Quaternary Research, 2014, 81(1): 151-157. doi: 10.1016/j.yqres.2013.10.001
[46] Iwasaki S, Takahashi K, Kanematsu Y, et al. Paleoproductivity and paleoceanography of the last 4.3 Myrs at IODP Expedition 323 Site U1341 in the Bering Sea based on biogenic opal content [J]. Deep Sea Research Part II:Topical Studies in Oceanography, 2016, 125-126: 145-154. doi: 10.1016/j.dsr2.2015.04.005
[47] Berger A. Orbital variations and insolation database[R]. Boulder CO, USA: National Oceanic and Atmospheric Administration, 1992.
-
期刊类型引用(2)
1. 何佶泳,田义超,张强,王栋华,张亚丽,周慧娟. 基于DEM的喀斯特峰丛洼地地貌信息提取及形态特征分析. 中国岩溶. 2024(03): 595-605 .  百度学术
百度学术
2. 赖荣贵. 机载雷达在藻砂分离监测中的应用研究. 测绘与空间地理信息. 2024(10): 182-184 . 百度学术
其他类型引用(3)
 下载:
下载:

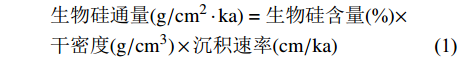
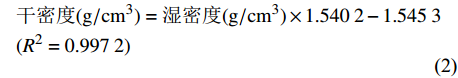
计量
- 文章访问数: 877
- HTML全文浏览量: 286
- PDF下载量: 80
- 被引次数: 5


