
Characteristics of organic carbon isotope and the paleoenvironmental significances of loess in Shengshan Island during the Last Glacial Period
-
摘要: 黄土沉积中有机碳同位素组成与古气候关系密切,对研究区域环境演变具有重要意义。本文聚焦我国东海嵊山岛末次冰期黄土地层,在磁化率和元素地球化学指标特征分析基础上,开展有机碳同位素组成特征研究。研究结果表明:嵊山岛黄土沉积剖面有机碳同位素组成波动范围−21.63‰~−27.56‰,平均值为−24.88‰,剖面有机碳同位素组成波动偏正;利用端元法对嵊山岛黄土沉积环境中C3/C4植被相对丰度进行估算,揭示沉积地层记录的植被类型是以C3型植被为主导,C4型植被对沉积地层中有机碳同位素的贡献有限;与南京洞穴石笋、苏禄海海洋沉积以及南极Vostok冰芯氧同位素记录对比,显示末次冰期间冰阶时期嵊山岛黄土沉积中有机碳同位素波动是以C3型植被为主导的植被碳同位素对降水条件改变的响应,降水可能是这一时期黄土沉积中有机碳同位素变化的主要影响因素;在末次冰消期阶段,温度有利于C4植被的发育,导致区域C4型植被丰度有所上升,并对沉积地层中有机碳同位素变动的贡献增加。Abstract: The organic carbon isotope composition in loess deposits is closely related to paleoclimate, which is of great significance to the study of regional environmental evolution. The loess deposit of the Last Glacial Period in Shengshan Island in the East China Sea off the East China was studied. The magnetic susceptibility and element geochemistry were analyzed, based on which the organic carbon isotope composition was scrutinized. Results show that since the Last Glaciation, the organic carbon isotope composition in the loess fluctuated from −21.63‰ to −27.56‰ on average of −24.88‰. In general, the value of organic carbon isotope decreased with the increase in burial depth of the loess. In addition, the relative abundance of C3/C4 vegetation in Shengshan Island was estimated by using the end-member method. It was revealed that the C3 plants dominated in the island since the Last Glacial Period, and C4 plants were very limited in the contribution to the organic carbon isotope in the loess deposit. In comparison with the oxygen isotope data of the cave stalagmites in Nanjing, marine sediments from Sulu Sea, and Antarctic Vostok ice core, the loess was deposited during the interstage of the Last Glaciation, and the fluctuation in organic carbon isotope value was resulted mainly from the responses of local ancient C3 plant-dominated vegetation to the variation of precipitation condition. The precipitation was the main influential factor on the organic carbon isotope variation during the period. In the last deglaciation stage, the paleo-temperature favored C4 plants booming, thus the relative abundance of C4 plants increased, and so did their contribution to the fluctuation of organic carbon isotopes in the loess deposits in the island.
-
中国黄土沉积分布范围广、时间跨度大,系统记录了亚洲季风系统变迁,成为开展古气候变化研究的重要载体[1-7]。黄土沉积的粒度组成、磁化率、元素地球化学和有机碳同位素等指标被广泛应用于古环境研究[8-16]。在有机碳同位素研究方面,由于C3和C4植物光合作用途径不同,使得植物碳同位素组成也不同,全球范围C3和C4植物碳同位素组成范围分别为–22‰~ –30‰和–9‰~ –19‰[17-19]。通过对黄土高原黄土研究发现,黄土沉积中有机碳主要来源于当地表生植被而非外源风尘输入,黄土沉积中有机碳同位素状况能够反映黄土堆积时期表生植被碳同位素组成情况[20-22],进而反演黄土堆积时期地表植被状况和古环境变化情况。相关研究显示,从末次冰盛期至全新世C4植被扩张,黄土高原黄土沉积中有机碳同位素记录该时期C4植被增加了约15%[23-26]。在黄土堆积的冰期时段,气候条件不利于C4植物发育,黄土沉积中有机碳同位素负偏;在古土壤发育的间冰期,温度升高、降水量增加,C4植被丰度增加,黄土沉积中有机碳同位素正偏[27-28],因此在冰期和间冰期时间尺度上,温度和降水等气候条件的变化影响黄土沉积中有机碳同位素组成。然而,不同区域黄土有机碳同位素变化对气候条件的响应也存在一定的差异性[29-31],也有研究显示末次冰期间冰阶时期黄土沉积中有机碳同位素负偏,此时黄土沉积中有机碳同位素组成的变化不指示C3和C4植被丰度变化,而是反映了以C3型植被为主导,植物碳同位素对温度、降水和二氧化碳浓度变化的综合响应,这也使得探讨黄土中有机碳同位素对气候变化的响应机制和过程变得复杂。嵊山岛黄土沉积位于东部季风区东缘,属典型风尘堆积,黄土沉积除接受西部内陆远源风尘物质外,东海陆架区域的近源风尘物质对黄土沉积也有贡献[32],对末次冰期东海岛屿黄土沉积的研究也是开展东部陆架区域古环境和古气候研究的重要内容,对认识东海岛屿所在区域末次冰期环境演变特征具有重要意义。本文以位于东海嵊山岛末次冰期黄土堆积地层为对象,通过黄土沉积地层样品中的有机碳同位素指标分析,同时结合磁化率和元素地球化学等代用指标,探讨黄土沉积有机碳同位素记录的区域古植被状况及其对古环境变化的响应,为东部海岸区域古植被和古气候重建提供支撑。
1. 区域概况
嵊山岛位于浙江沿海舟山群岛外缘区域,处于东海马鞍列岛核心位置,隶属舟山市嵊泗县(图1)。嵊山岛岛屿面积约4.22 km2,海岸线总长度约19.26 km,地貌类型多低山丘陵,气候属北亚热带海洋季风气候,受海洋气流作用明显,夏季盛行东南风,高温多雨;冬季盛行西北风,低温干燥,年均气温约16.4 °C,平均降水量1072 mm。嵊山岛属嵊泗列岛北部区域,该区域广泛分布落叶阔叶林,落叶树种以天仙果、白栎、日本野桐、黄檀和山槐为主,乔木林高度普遍低矮,无人小岛大多被灌木和草丛覆盖[33]。嵊山岛主要由花岗岩组成,大部分区域基岩直接出露地表,在岛屿的山坡、山谷以及低洼地带形成第四纪松散堆积物,片状黄土覆盖在基岩山坡上,并随岛屿地势起伏而变化[32]。
本文研究所选黄土剖面位于嵊山岛陈钱山东北坡(30.73°N、122.817°E),海拔约150 m。剖面整体呈黄棕色和黄褐色,沉积地层质地坚硬、结构致密,垂直节理发育并伴有少量钙质结核,整个沉积剖面黄土颗粒较细,无明显层理,沉积剖面上未见花岗岩风化残留物。剖面底部为花岗岩基岩风化壳,部分花岗岩风化颗粒与黄土沉积相混合,黄土沉积剖面底部与该花岗岩风化壳呈不整合接触,顶部土质较为疏松,受人为活动影响较大,整个剖面深度约280 cm。为获取沉积地层古环境信息,同时考虑到人为活动影响,由地表以下5~275 cm剖面深度以3 cm为间隔获取黄土沉积样品。
2. 研究方法
2.1 地层年代测试
在沉积剖面10、100、150和250 cm深度采集4个光释光样品,在暗室红光条件下完成样品前处理,取出采集的样品,除去两端可能曝光部分,剩余样品依次使用30%的双氧水和10%的盐酸去除样品中有机质和碳酸盐,经过湿筛和去除重矿物,以及氢氟酸刻蚀等步骤,获得测年所需石英颗粒。采样管两端样品用于U、Th、K以及含水量指标测定,利用单片再生剂量法(single aliquot regenerative-dose,SAR)完成所提取石英等效剂量的测定。样品含水量为实测含水量,样品的U、Th、K含量以中子活化法(NAA)测试,样品光释光年代测量在南京大学地理与海洋科学学院光释光测年实验室完成。
2.2 总有机碳和有机碳同位素分析
称取约 2 g 黄土样品置于80 mL的聚丙烯离心管中,加入20 mL浓度为2 mol/L 的盐酸,充分摇匀,静置24 h,期间摇匀样品2—3 次,使其充分反应。24 h 后,倒出上清液,加入超纯水,离心,倒出上清液,重复上述水洗步骤(本次实验6—7次)直至pH试纸检测上清液显示中性。然后将样品置于烘箱中烘干(温度设置在45℃),借助研钵将样品研磨至均质化,将样品干燥保存待用。有机碳使用Vario EL元素分析仪测定,分析精度±2%,有机碳同位素使用DELTA Plus XL稳定同位素质谱仪(Finnigan MAT 公司)测定,分析精度为±0.1‰,样品测试工作在华东师范大学河口海岸学国家重点实验室完成。
2.3 地层磁化率测定
将所采集样品置于 35℃的烘箱内恒温烘干。称取7 g 烘干的样品用塑料保鲜膜包起来,装入体积为10 mL 的磁学专用塑料盒中,使用华东师范大学河口海岸学国家重点实验室的Bartington (英国制造)双频磁化率仪MS2 进行磁化率的测定。
2.4 地球化学元素测试
在地球化学元素分析过程中,将黄土样品置于玛瑙研磨碗中研磨,并过50 μm筛。称取约5 g研磨后的过筛样品倒入直径约4 cm、厚约0.5 cm的圆形模具中央,将样品送至压样机压制。压样机加压并保持在37.5 t/m2约30 s后,自然泄压,待测样品制备完成。将制作好的样品置于X荧光光谱仪(XRF)(日本)中进行测试,完成所需Rb、Sr元素测定,测试过程中使用GSD-9标准物质进行质量控制,样品分析误差小于5%。
3. 结果和讨论
3.1 年代结果
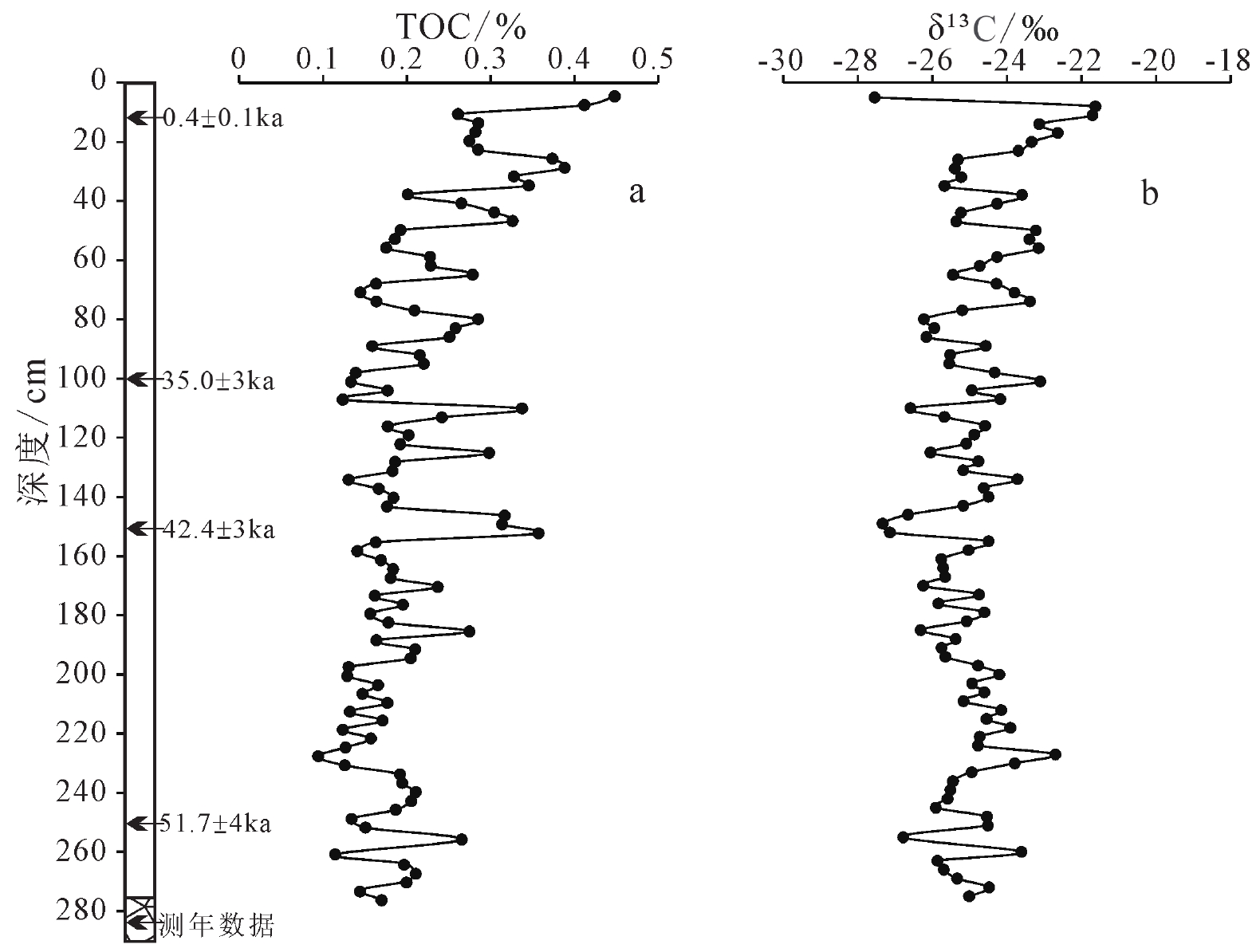
如表1所示,嵊山岛黄土沉积在剖面深度10、100、150和250 cm处光释光年代测试结果分别为0.4±0.1、35.03±3、42.41±4和51.67±4 ka。嵊山岛黄土沉积的采样剖面质地均一,未发现地层倒置情况,结合光释光年代测定结果,嵊山岛黄土堆积于末次冰期。
表 1 嵊山岛黄土剖面光释光年代测定结果Table 1. OSL (optically stimulated luminescence) dating results of the loess in Shengshan Island深度/cm U/(μg/g) Th/(μg/g) K/% 实测含水量/% 环境剂量率/(Gy/ka) 等效剂量/Gy 年龄/ka 10 2.83 14.3 1.49 14.45 3.05 1.2±0.3 0.4±0.1 100 2.98 15.1 1.69 11.86 3.34 117±4 35.03±3 150 2.94 14.4 1.56 8.18 3.23 137±8 42.41±4 250 2.93 14.3 1.62 16.49 3.00 152±5 51.67±4 3.2 总有机碳(TOC)和有机碳同位素特征
嵊山岛黄土沉积剖面有机碳(TOC)含量整体较低(图2a),含量范围0.09%~0.45%,平均值为0.21%,变化幅度为0.36%。总体而言,随嵊山岛黄土沉积剖面深度的减小,总有机碳含量呈现波动上升的变化趋势,从剖面底部至80 cm深度附近,总有机碳含量整体较低,波动明显,剖面平均值为0.19%;从剖面80 cm深度附近至地表,总有机碳波动明显,总体呈上升趋势,平均含量达0.27%,地表附近总有机碳含量最高,达0.45%,地表腐殖质堆积淋溶可能是近地表总有机碳含量上升的主要原因。现代植被研究显示,C3型植物 δ13C 组成多集中于 −27‰,分布范围 −20‰~−34‰,C4 型植物 δ13C组成多集中于−13‰,分布范围–9‰~–19‰[17-18, 34]。土壤中δ13C大于–14‰代表纯C4型植被,−14‰~−19‰之间表示C4型植被为优势植被,在−19‰~−24‰之间表示C3型植被为优势植被,小于−24‰可代表纯C3型植被[35]。对美国大平原和澳大利亚现代土壤中有机碳同位素研究显示,当土壤有机碳小于−24‰时,地表植被为纯C3型植物[36-37],通过对中国东部表土有机碳同位素研究,发现我国31°N以南的东部地区C3型植物占主导地位[38]。
![]() 图 2 嵊山岛黄土剖面总有机碳含量(a)和有机碳同位素组成特征(b)Figure 2. Characteristics of TOC (total organic carbon) content (a) and δ13C composition (b) in the Shengshan Island loess
图 2 嵊山岛黄土剖面总有机碳含量(a)和有机碳同位素组成特征(b)Figure 2. Characteristics of TOC (total organic carbon) content (a) and δ13C composition (b) in the Shengshan Island loess土壤表生植物残体的分解累积成为土壤有机质的主要来源,通过对土壤中脂类化合物烷烃同位素分析发现,土壤中有机质主要来源于上覆植被,因此土壤中有机碳同位素组成继承了上覆植被的碳同位素特征[20,39]。通过对风成黄土的研究显示,黄土-古土壤层中有机质主要来源于地表植被,外源风尘输入对黄土沉积中有机碳影响十分有限,黄土沉积中的有机碳同位素组成的变化可以反映黄土堆积时期表生植被演化[22,40]。嵊山岛黄土沉积剖面有机碳同位素δ13C组成变化范围为−21.63‰~−27.56‰(图2b),平均值为−24.88‰,变化幅度为5.93‰。所测嵊山岛黄土沉积剖面δ13C值波动明显,多数样品都小于−24‰,这也显示末次冰期该研究区域植被类型可能是以C3型植物为主导。
3.3 有机碳同位素组成与C4植物丰度估算
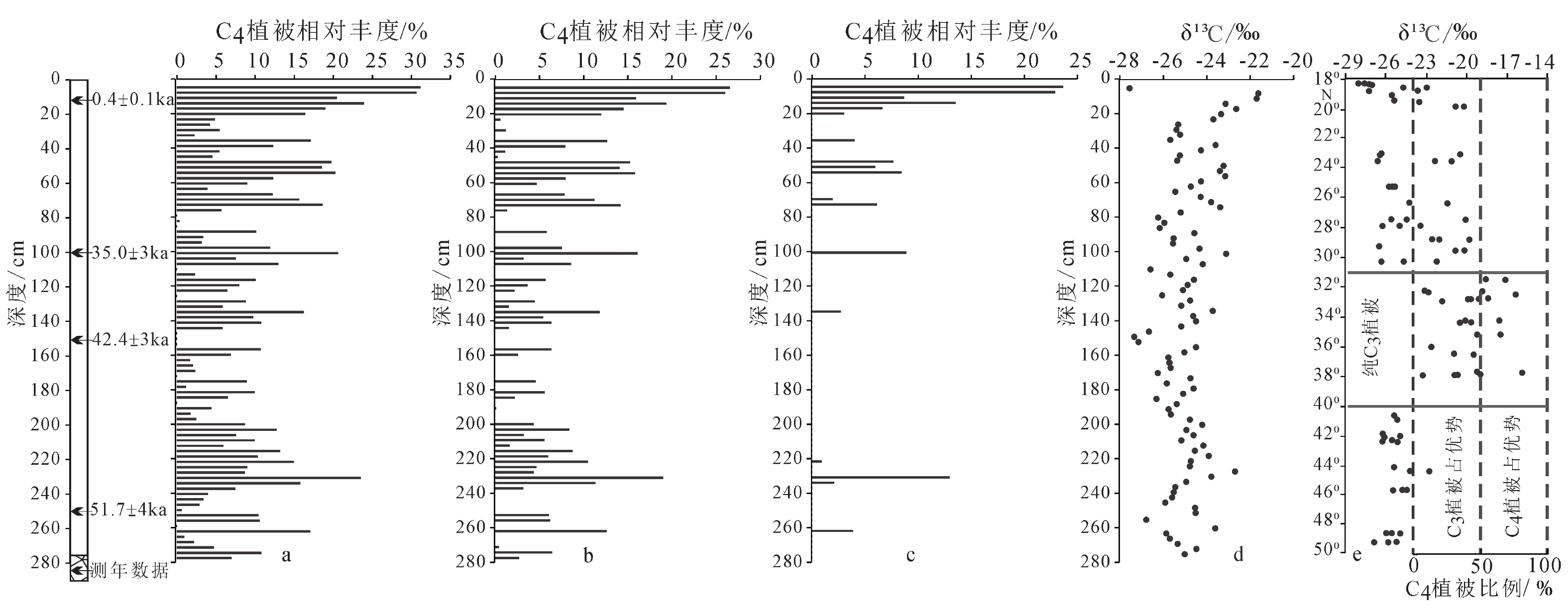
土壤中有机碳同位素组成受到当地原生植被影响,可以反映植被组成变化[41]。对黄土高原黄土沉积研究显示黄土中有机碳同位素主要来源于当地植被,有机碳同位素组成可以指示相应时期植被碳同位素状况,进而反演当地古植被的演替[22]。在古植被的定量估算研究中,可以使用二端元混合模型δ13CTOC=Fδ13C4+(1−F)δ13C3对C3/C4植被生物量的相对比例进行估算[23,42]。其中δ13CTOC表示所测土壤中有机碳同位素组成,δ13C3和δ13C4分别表示C3和C4植被类型的端元碳同位素组成,F表示C4型植物生物量在植被中的相对占比,指示C4型植被相对丰度。通过二端元混合模型计算C4植物生物量占比,需要确定C3和C4植被类型的碳同位素端元值。由于土壤中有机碳同位素组成受植被平均碳同位素组成的影响,已知全球范围内C3型植被δ13C3平均值为−27‰,C4型植被δ13C4平均值为−13‰,相关研究以此作为碳同位素端元值,开展C4植物生物量占比估算[28]。通过二端元混合模型估算C4植物生物量占比,在选择植物有机碳同位素端元值过程中还需考虑植物有机质在转化为土壤有机质过程中的同位素分馏作用。由于同位素分馏作用,土壤有机质中碳同位素要比对应植物碳同位素偏重,土壤有机质与对应植被之间产生1‰或 1.3‰ 的碳同位素分馏作用,也可在此基础上确定土壤有机碳同位素端元值开展C3/C4植被丰度计算 [43-44]。也有研究根据美国大平原和澳大利亚现代表土有机碳同位素与上覆植被情况,选择 −24‰ 和−14‰分别作为纯C3和C4植被下土壤中有机碳同位素端元值,对黄土高原地区古植被情况开展研究,并取得相应研究成果[26]。在开展中更新世以来东部季风区哈尔滨黄土有机碳同位素研究中,结合区域现代C3和C4植被有机碳同位素组成,并考虑1.3‰ 的碳同位素分馏效应,分别选取 −25.4‰ 和−11.2‰作为两种植被对应土壤有机碳同位素端元值,开展C4植被丰度估算[44-45]。为更加全面地分析末次冰期嵊山岛黄土沉积区域植被构成,分别通过选取−25.4‰和−11.2‰、−24‰和−14‰作为碳同位素端元值进行植被丰度计算,同时选择C3、C4植物碳同位素组成全球平均值,并在考虑土壤有机质与对应植被碳同位素1‰分馏作用基础上,以−26‰和−12‰作为C3和C4型植被对应的土壤有机碳同位素端元值,分别开展C4型植被相对丰度估算(图3)。
![]() 图 3 嵊山岛黄土沉积中C4植被相对丰度估算a: 参考全球植物碳同位素组成[28],b: 参考哈尔滨黄土所选端元[44-45],c: 参考黄土高原黄土所选端元[26],d: 嵊山岛黄土有机碳同位素分布,e: 中国东部C3/C4植被丰度变化[38]。Figure 3. The calculated relative abundance of C4 plants in the Shengshan Island loessa: The endmember value considering global plant carbon isotope composition[28], b: the endmember value considering Harbin loess deposit[44-45], c: the endmember value considering the Loess Plateau loess deposit[26], d: profile of organic carbon isotope in Shengshan Island, e: variations of C3 and C4 abundance in the eastern China[38].
图 3 嵊山岛黄土沉积中C4植被相对丰度估算a: 参考全球植物碳同位素组成[28],b: 参考哈尔滨黄土所选端元[44-45],c: 参考黄土高原黄土所选端元[26],d: 嵊山岛黄土有机碳同位素分布,e: 中国东部C3/C4植被丰度变化[38]。Figure 3. The calculated relative abundance of C4 plants in the Shengshan Island loessa: The endmember value considering global plant carbon isotope composition[28], b: the endmember value considering Harbin loess deposit[44-45], c: the endmember value considering the Loess Plateau loess deposit[26], d: profile of organic carbon isotope in Shengshan Island, e: variations of C3 and C4 abundance in the eastern China[38].结合中国东部表土有机碳同位素数据,对中国东部地区C4植物相对生物量贡献进行估算,中国东部31°~40°N区域以南为纯C3植物或者以C3植物占优势的C3/C4混合植被[38]。通过对湖泊沉积中的碳同位素研究,显示我国南方热带-亚热带区域冰期中以C3型植被为主导[46]。利用3组不同端元值,对末次冰期嵊山岛黄土沉积剖面C4植物相对丰度进行估算显示:以全球植物碳同位素平均组成为端元值并考虑碳同位素1‰分馏作用基础上,估算嵊山岛黄土沉积剖面C4植物相对丰度最大值为31.2%,平均值为8.5%;以东部季风区哈尔滨黄土根据区域C3和C4植被有机碳同位素组成所选取的端元值进行估算,显示剖面C4植物相对丰度最大值为26.5%,平均值为5.1%;以黄土高原等地研究中统一所选择的–24‰和–14‰作为碳同位素端元值进行估算,显示剖面C4植物相对丰度最大值为23.7%,平均值为1.7%。由于没有对研究区域内现代植被碳同位素组成进行调查研究,同时考虑现代植被的碳同位素组成在不同的气候条件下可能也不相同,为了更加全面地判断末次冰期嵊山岛区域古植被的变化趋势,选取多组端元值对植被丰度进行估算。其中,参照北美大平原和澳大利亚表土有机碳同位素研究结果,选取−24‰和−14‰作为纯C3和C4型植被下土壤有机碳同位素端元值,估算黄土高原末次冰期至全新世C3、C4型植被丰度变化,取得了较好的研究成果[47],因此文中也将使用该端元值估算结果进行后续的讨论分析。通过选取的不同端元值分别对嵊山岛黄土沉积剖面C4植被丰度进行估算,均显示末次冰期地处东部季风区的嵊山岛黄土沉积区域C4植被占比低,而C3植被始终占据绝对优势,这与通过表土所估算的我国东部环北太平洋相应区域现代植被中C3/C4植物相对丰度大体一致[38]。
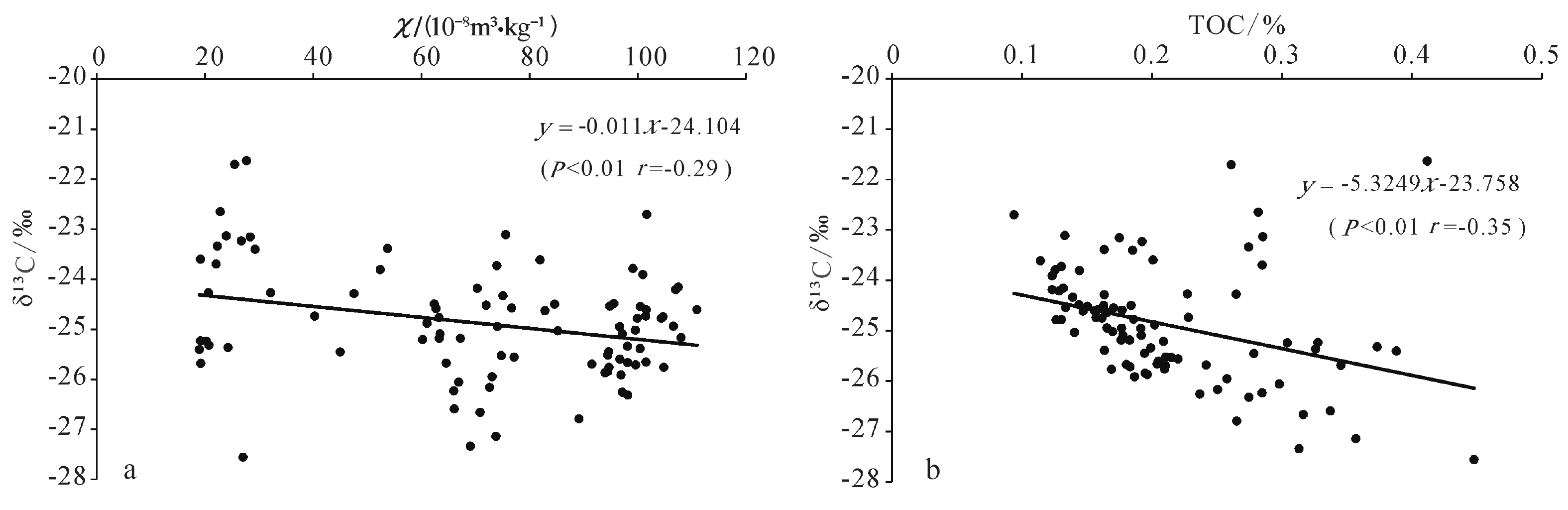
已有研究显示,在黄土高原东部地区古土壤形成阶段有机碳同位素偏正,黄土层中有机碳同位素偏负,黄土沉积中总有机碳与有机碳同位素正相关,有机碳同位素可以揭示C3和C4植被相对丰度变化[23]。但是在黄土高原西南部,古土壤形成阶段有机碳同位素偏负,有机碳同位素与总有机碳呈负相关,揭示以C3型植被为主导的植被类型对温度和降水等气候条件的综合响应[29]。通过对现代C3 型植物研究发现,C3型植物碳同位素组成与降水量负相关,我国北方黄土区年降水量每增加100 mm, C3植被有机碳同位素平均值偏负1.1‰左右[48-51]。对我国降水量400 mm的半干旱地区C3植被碳同位素研究显示,温度每上升1 °C,C3植物碳同位素平均值偏重0.104‰[52]。黄土磁化率变化可以反映降水量情况,指示季风强度变化[53],与有机碳同位素变化在冰期和间冰期尺度上具有一定相关性[54]。通过分析嵊山岛黄土沉积剖面末次冰期有机碳同位素与磁化率和总有机碳的相关关系(图4),发现嵊山岛黄土沉积剖面有机碳同位素组成与剖面磁化率和总有机碳均显示负相关的趋势。
![]() 图 4 嵊山岛黄土沉积剖面有机碳同位素与磁化率(a)和总有机碳散点图(b)Figure 4. Scatter plots of the relationships between δ13C and magnetic susceptibility (a), and between δ13C and TOC (b) of the Shengshan Island loess
图 4 嵊山岛黄土沉积剖面有机碳同位素与磁化率(a)和总有机碳散点图(b)Figure 4. Scatter plots of the relationships between δ13C and magnetic susceptibility (a), and between δ13C and TOC (b) of the Shengshan Island loess进一步分析,嵊山岛黄土沉积剖面中有机碳同位素与磁化率呈负相关趋势(图4a),在P<0.01水平下,相关系数为–0.29;与总有机碳同样呈现负相关趋势(图4b);P<0.01水平下,相关系数为–0.35。说明在末次冰期间冰阶时期,东部季风区的夏季风增强时段,黄土沉积磁化率数值升高,气候相对湿润。由于区域以C3型植被为主导,δ13C负偏,间接指示了较为湿润的气候条件,气候湿润有利于植被发育,进而产生较高的生物量,表生植被生物量的增加使得黄土沉积中有机质含量上升,进而使得总有机碳含量增加。嵊山岛区域末次冰期黄土沉积磁化率升高,有机碳同位素负偏,间接反映了夏季风条件的改善。黄土沉积有机碳同位素与磁化率指标的负相关关系,也进一步揭示了黄土堆积时期以C3型植被为主导的植物碳同位素变化对气候条件的响应。
3.4 嵊山岛黄土沉积有机碳同位素对气候条件的响应
大气二氧化碳浓度、温度和降水量影响陆生植被及其碳同位素组成[55],大气中二氧化碳浓度降低,温度增加,以及相对干旱的气候条件,有利于C4植被发育[34],反之将促进C3植被发育。嵊山岛黄土剖面有机碳同位素组成显示末次冰期该区域C3型植被占绝对优势地位,黄土沉积中有机碳同位素受区域内C3型植被碳同位素组成影响。相关研究显示,大气中二氧化碳浓度每降低100×10−6,C3型植物碳同位素偏正2.0‰±0.1‰[56],末次冰期大气中二氧化碳浓度的变化幅度约30×10−6 [29],这样末次冰期因二氧化碳浓度降低,使得黄土沉积中有机碳同位素偏正约0.67‰±0.033‰,取最大值偏正约0.70‰。同时,温度每上升1°C,C3植物碳同位素平均偏重0.104‰[52],末次冰期间冰阶至末次盛冰期全球温度变幅约5 °C[57],大陆中低纬度的低海拔地区降温约6 °C[58],取温度变幅平均值5.5 °C计算,末次冰期至末次盛冰期通过C3型植物碳同位素响应,影响黄土沉积中有机碳同位素偏轻约0.57‰,因此嵊山岛黄土沉积从末次冰期间冰阶至末次盛冰期,全球二氧化碳浓度和温度降低的综合影响为正偏0.13‰,该时段全球二氧化碳浓度和温度降低幅度对黄土沉积中有机碳同位素组成影响较小。基于对现代C3型植被研究,降水量的增加与C3型植被碳同位素组成呈负相关关系[48-50],降水量每增加100 mm, C3型植被碳同位素负偏1.1‰[51],末次冰期间冰阶比末次盛冰期降水量多250~310 mm[29],计算末次冰期降水量变化影响黄土沉积中有机碳同位素变幅为2.75‰~3.41‰。末次冰期以来嵊山岛黄土剖面记录有机碳同位素变幅大于5‰,末次冰期间冰阶至末次盛冰期,嵊山岛黄土沉积中有机碳同位素偏正约3.63‰。剖面有机碳同位素波动可能主要受降水量的影响,降水量的变化可能是影响末次冰期嵊山岛黄土沉积中有机碳同位素组成波动的主要原因。
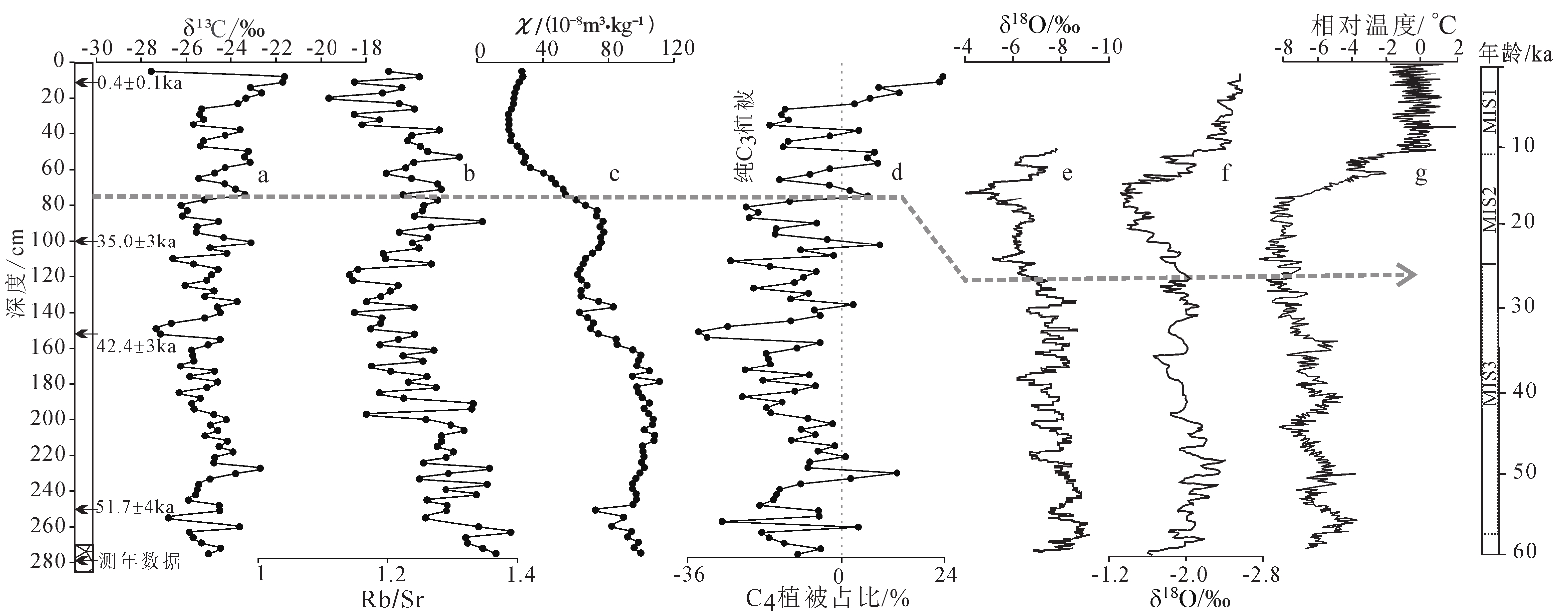
铷锶元素两者的活动性不同,具有不同的地球化学行为,铷锶分别作为稳定性和活动性组分,两者比值可以反映风化成壤强度,指示黄土沉积所经历的暖湿程度,进而记录夏季风环流强度的变化[59]。黄土沉积中磁性矿物种类、含量以及磁性颗粒的大小蕴含着丰富的古环境信息,黄土沉积中磁化率指标与黄土堆积时期的降水和温度密切相关,磁化率数值的高低也可以指示夏季风强弱的变化[2,60]。黄土有机碳同位素主要受温度和季风作用下的季节性降水影响,冰期气候条件不利于C4植被发育,C4植被丰度降低[23]。对末次冰期陇西黄土高原塬堡剖面研究显示,有机碳同位素组成的变化主要是C3植被本身对温度、降水等条件的响应,黄土沉积中有机碳同位素负偏与夏季风增强和降水增加时段相吻合,黄土剖面末次冰期有机碳同位素变化主要受控于降水条件的变化[29]。对西北欧黄土中有机碳同位素分析发现,末次冰期以C3型植物占据优势,黄土中有机碳同位素的变化反映了C3植物本身碳同位素组成对降水条件和二氧化碳浓度变化的响应,有机碳同位素可以作为指示降水量变化的潜在代用指标[31,61]。根据嵊山岛黄土沉积剖面中有机碳同位素数值波动,将沉积剖面划分为2个阶段(图5):第一阶段,从剖面底部至75 cm深度附近,大体对应深海氧同位素MIS3阶段。该沉积阶段黄土沉积剖面磁化率在高值区域波动,平均值为86.56×10−8 m3/kg,Rr/Sr比值先波动降低然后波动上升,平均值为1.25;洞穴石笋和海洋沉积中氧同位素记录δ18O在偏负区域波动。剖面磁化率、铷锶比值指示嵊山岛黄土剖面在MIS3阶段经历相对湿润和温暖的沉积环境。根据Vostok冰芯记录该沉积时段温度状况相对于现代大约低6℃,相对温度在该沉积阶段集中于−6℃附近波动,石笋和深海氧同位素记录偏负,同样指示该沉积时段相对暖湿的气候环境。此时段嵊山岛黄土沉积剖面中δ13C数值大都在小于−24‰一侧波动,该沉积阶段环境植被基本为C3型植被,剖面δ13C值的相对偏负,指示以C3型植被为主导的区域植被碳同位素对降水条件的响应,进而剖面δ13C值的波动可以指示降水条件的变化。第二阶段,剖面75~25 cm附近,大体对应深海氧同位素MIS2阶段。该沉积阶段剖面磁化率数值先下降后略回升,Rr/Sr比值先波动降低后波动上升。地处中国东部季风区的石笋氧同位素δ18O记录及深海氧同位素δ18O记录,较上一阶段均显示先波动偏正然后波动偏负,指示温度和降水量先降低而后增加。在温度变化方面,Vostok冰芯记录该沉积时段相对温度状况较上一沉积阶段先进一步波动降低,然后再快速回升。在末次盛冰期阶段,相对温度集中在–8℃附近波动[64],进入冰消期后温度快速波动上升。该阶段嵊山岛黄土沉积剖面中部分δ13C数值在大于−24%一侧波动,黄土沉积中δ13C数值较上一阶段波动偏正。大气中二氧化碳浓度升高和降水条件改善,C3型植物碳同位素组成偏负[48-50,56],末次冰消期至全新世过渡过程中,在二氧化碳浓度上升、降水增加条件下,黄土沉积剖面δ13C数值相对偏正,揭示区域环境中C4型植被发育,通过估算,该阶段部分沉积时段C4植被丰度达15%左右。与末次冰期相比,末次冰消期温度条件大幅改善,温度相对于末次冰期上升8~10℃[61],对末次冰期以来陆地植被中C3/C4植被相对丰度研究显示,在末次冰期以来的大气二氧化碳浓度背景下,温度成为中纬度地区末次冰期至全新世阶段C4植被丰度上升的首要控制因素[65],温度达到C4植物生长“阈值”,对C4植物生长起到促进作用[24]。嵊山岛黄土在末次盛冰期至冰消期沉积阶段,黄土剖面有机碳同位素δ13C数值波动响应了因温度变化而主导的C4植被丰度变化,显示出该区域在以C3型植物主导下,因温度的上升促使 C4植被发育的过程。此时段沉积剖面有机碳同位素波动更多是对温度变化响应的结果。
![]() 图 5 嵊山岛黄土沉积剖面有机碳同位素、铷锶比、磁化率、C4植被丰度变化a: 嵊山岛黄土沉积有机碳同位素组成, b: 嵊山岛黄土沉积铷锶比, c: 嵊山岛黄土沉积磁化率, d: 嵊山岛黄土沉积C4植被丰度, e: 南京葫芦洞石笋中氧同位素记录[62],f: Sulu Sea 氧同位素记录[63] , g: Vostok冰芯记录相对温度变化[64]。Figure 5. Profiles of δ13C, Rb/Sr, magnetic susceptibility (χ), and relative C4 vegetation abundance in the Shengshan Island loessa: δ13C composition, b: Rb/Sr, c: magnetic susceptibility (χ) , d: the abundance of C4 vegetation, e: oxygen-isotope (δ18O) records of the stalagmites in Hulu Cave, Nanjing[62], f: oxygen-isotope (δ18O) records in Sulu Sea[63], g: relative temperature changes shown in the Antarctic Vostok ice cores [64].
图 5 嵊山岛黄土沉积剖面有机碳同位素、铷锶比、磁化率、C4植被丰度变化a: 嵊山岛黄土沉积有机碳同位素组成, b: 嵊山岛黄土沉积铷锶比, c: 嵊山岛黄土沉积磁化率, d: 嵊山岛黄土沉积C4植被丰度, e: 南京葫芦洞石笋中氧同位素记录[62],f: Sulu Sea 氧同位素记录[63] , g: Vostok冰芯记录相对温度变化[64]。Figure 5. Profiles of δ13C, Rb/Sr, magnetic susceptibility (χ), and relative C4 vegetation abundance in the Shengshan Island loessa: δ13C composition, b: Rb/Sr, c: magnetic susceptibility (χ) , d: the abundance of C4 vegetation, e: oxygen-isotope (δ18O) records of the stalagmites in Hulu Cave, Nanjing[62], f: oxygen-isotope (δ18O) records in Sulu Sea[63], g: relative temperature changes shown in the Antarctic Vostok ice cores [64].4. 结论
(1)末次冰期嵊山岛黄土沉积剖面有机碳同位素组成波动明显,剖面有机碳同位素δ13C数值大都在小于–24‰范围波动,有机碳同位素组成范围为–21.63‰~ –27.56‰,平均值为–24.88‰。利用二端元混合模型对末次冰期嵊山岛黄土沉积环境中C3/C4植被相对比例进行估算,揭示末次冰期嵊山岛黄土沉积时段的植被类型以C3型植被占绝对优势,末次盛冰期至末次冰消期C4植被丰度有所上升。
(2)通过与洞穴石笋、南极冰芯和海洋沉积氧同位素组成的对比,显示末次冰期嵊山岛黄土沉积中有机碳同位素δ13C数值波动主要受到降水条件的影响,降水条件是影响末次冰期间冰阶黄土沉积剖面有机碳同位素负偏的主要因素,末次冰期间冰阶黄土沉积中有机碳同位素波动主要指示C3植被碳同位素对降水条件的响应,黄土沉积有机碳同位素组成的波动反映降水条件变化。从末次盛冰期至末次冰消期的黄土沉积阶段,温度快速回升并达到C4植被发育“阈值”,C4植被生长。此时,黄土沉积剖面中δ13C数值波动偏正可能主要是对温度条件改变的响应,沉积剖面中有机碳同位素δ13C数值波动可能揭示了温度主导下的C4植被丰度变化,温度成为该时期碳同位素组成变化的主要影响因素。
-
![]()
图 2 嵊山岛黄土剖面总有机碳含量(a)和有机碳同位素组成特征(b)
Figure 2. Characteristics of TOC (total organic carbon) content (a) and δ13C composition (b) in the Shengshan Island loess
![]()
图 3 嵊山岛黄土沉积中C4植被相对丰度估算
a: 参考全球植物碳同位素组成[28],b: 参考哈尔滨黄土所选端元[44-45],c: 参考黄土高原黄土所选端元[26],d: 嵊山岛黄土有机碳同位素分布,e: 中国东部C3/C4植被丰度变化[38]。
Figure 3. The calculated relative abundance of C4 plants in the Shengshan Island loess
a: The endmember value considering global plant carbon isotope composition[28], b: the endmember value considering Harbin loess deposit[44-45], c: the endmember value considering the Loess Plateau loess deposit[26], d: profile of organic carbon isotope in Shengshan Island, e: variations of C3 and C4 abundance in the eastern China[38].
![]()
图 4 嵊山岛黄土沉积剖面有机碳同位素与磁化率(a)和总有机碳散点图(b)
Figure 4. Scatter plots of the relationships between δ13C and magnetic susceptibility (a), and between δ13C and TOC (b) of the Shengshan Island loess
![]()
图 5 嵊山岛黄土沉积剖面有机碳同位素、铷锶比、磁化率、C4植被丰度变化
a: 嵊山岛黄土沉积有机碳同位素组成, b: 嵊山岛黄土沉积铷锶比, c: 嵊山岛黄土沉积磁化率, d: 嵊山岛黄土沉积C4植被丰度, e: 南京葫芦洞石笋中氧同位素记录[62],f: Sulu Sea 氧同位素记录[63] , g: Vostok冰芯记录相对温度变化[64]。
Figure 5. Profiles of δ13C, Rb/Sr, magnetic susceptibility (χ), and relative C4 vegetation abundance in the Shengshan Island loess
a: δ13C composition, b: Rb/Sr, c: magnetic susceptibility (χ) , d: the abundance of C4 vegetation, e: oxygen-isotope (δ18O) records of the stalagmites in Hulu Cave, Nanjing[62], f: oxygen-isotope (δ18O) records in Sulu Sea[63], g: relative temperature changes shown in the Antarctic Vostok ice cores [64].
表 1 嵊山岛黄土剖面光释光年代测定结果
Table 1 OSL (optically stimulated luminescence) dating results of the loess in Shengshan Island
深度/cm U/(μg/g) Th/(μg/g) K/% 实测含水量/% 环境剂量率/(Gy/ka) 等效剂量/Gy 年龄/ka 10 2.83 14.3 1.49 14.45 3.05 1.2±0.3 0.4±0.1 100 2.98 15.1 1.69 11.86 3.34 117±4 35.03±3 150 2.94 14.4 1.56 8.18 3.23 137±8 42.41±4 250 2.93 14.3 1.62 16.49 3.00 152±5 51.67±4  下载: 导出CSV
下载: 导出CSV
-
[1] 刘东生. 黄土与环境[M]. 北京: 科学出版社, 1985 LIU Tungsheng. Loess and the Environment[M]. Beijing: Science Press, 1985.
[2] Maher B A. Palaeoclimatic records of the loess/palaeosol sequences of the Chinese Loess Plateau [J]. Quaternary Science Reviews, 2016, 154: 23-84. doi: 10.1016/j.quascirev.2016.08.004
[3] Yang H, Li G Q, Gou S Y et al. The close-space luminescence dated loess record from SW Junggar Basin indicates persistent aridity during the last glacial-interglacial cycle in lowlands of Central Asia [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2021, 584: 110664. doi: 10.1016/j.palaeo.2021.110664
[4] Li P, Zhang C X, Wu H B et al. Geochemical characteristics of Holocene loess-paleoslol sequences in central Chinese Loess Plateau and their implications for East Asian monsoon evolution [J]. Quaternary International, 2022, 616: 99-108. doi: 10.1016/j.quaint.2021.10.017
[5] Xu X W, Qiang X K, Hu S et al. Records of the Mid-Brunhes Event in Chinese loess-paleosol sequences [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 543: 109596. doi: 10.1016/j.palaeo.2020.109596
[6] Mir J A, Dar R A, Vinnepand M et al. Environmental reconstruction potentials of loess-paleosol-sequences in Kashmir through high-resolution proxy data [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2022, 601: 111100. doi: 10.1016/j.palaeo.2022.111100
[7] Wang Y, Guo F, Ma L et al. Millennial-scale summer monsoon oscillations over the last 260 ka revealed by high-resolution elemental results of the Mangshan loess-palaeosol sequence from the southeastern Chinese Loess Plateau [J]. Quaternary International, 2020, 552: 164-174. doi: 10.1016/j.quaint.2020.05.039
[8] 周家兴, 于娟, 杨丽君, 等. 铜川地区早中全新世黄土沉积特征及其古气候意义[J]. 海洋地质与第四纪地质, 2020, 40(1):160-166 ZHOU Jiaxing, YU Juan, YANG Lijun et al. Sedimentary characteristics of the early and middle Holocene loess in Tongchuan area and their implications for paleoclimate [J]. Marine Geology & Quaternary Geology, 2020, 40(1): 160-166.
[9] Ding Z L, Yu Z W, Yang S L et al. Coeval changes in grain size and sedimentation rate of eolian loess, the Chinese Loess Plateau [J]. Geophysical Research Letters, 2001, 28(10): 2097-2100. doi: 10.1029/2000GL006110
[10] Kong X H, Zhou W J, Beck J W et al. Loess magnetic susceptibility flux: A new proxy of East Asian monsoon precipitation [J]. Journal of Asian Earth Sciences, 2020, 201: 104489. doi: 10.1016/j.jseaes.2020.104489
[11] 刘秀铭, 刘东生, Heller F, 等. 黄土频率磁化率与古气候冷暖变换[J]. 第四纪研究, 1990,10(1):42-50 doi: 10.3321/j.issn:1001-7410.1990.01.005 LIU Xiuming, LIU Tungsheng, Heller F et al. Frequency-dependent susceptibility of loess and quaternary paleoclimate [J]. Quaternary Sciences, 1990,10(1): 42-50. doi: 10.3321/j.issn:1001-7410.1990.01.005
[12] 石 浩, 岳大鹏, 赵景波, 等. 陕西绥德地区黄土-古土壤序列地球化学特征及其环境指示意义[J]. 地球与环境, 2022, 50(1):1-13 SHI Hao, YUE Dapeng, ZHAO Jingbo et al. Geochemical characteristics of loess paleosol sequence and its environmental implications in Suide area, Shanxi [J]. Earth and Environment, 2022, 50(1): 1-13.
[13] 田庆春, 郝晓龙, 韩军青, 等. 临汾盆地黄土沉积微量元素地球化学特征及其气候意义[J]. 干旱区资源与环境, 2022, 36(5):87-93 doi: 10.13448/j.cnki.jalre.2022.123 TIAN Qingchun, HAO Xiaolong, HAN Junqing et al. Geochemical characteristics and climatic significance of trace elements in loess of Linfen basin [J]. Journal of Arid Land Resources and Environment, 2022, 36(5): 87-93. doi: 10.13448/j.cnki.jalre.2022.123
[14] Yang S L, Liu L, Chen H et al. Variability and environmental significance of organic carbon isotopes in Ganzi loess since the last interglacial on the eastern Tibetan Plateau [J]. Catena, 2021, 196: 104866. doi: 10.1016/j.catena.2020.104866
[15] Zhou B, Wali G, Peterse F et al. Organic carbon isotope and molecular fossil records of vegetation evolution in central Loess Plateau since 450 kyr [J]. Science China Earth Sciences, 2016, 59(6): 1206-1215. doi: 10.1007/s11430-016-5276-x
[16] An Z S, Huang Y S, Liu W G et al. Multiple expansions of C4 plant biomass in East Asia since 7 Ma coupled with strengthened monsoon circulation [J]. Geology, 2005, 33(9): 705-708. doi: 10.1130/G21423.1
[17] O’Leary M H. Carbon isotope fractionation in plants [J]. Phytochemistry, 1981, 20(4): 553-567. doi: 10.1016/0031-9422(81)85134-5
[18] O’Leary M H. Carbon isotope in photosynthesis [J]. BioScience, 1988, 38(5): 328-336. doi: 10.2307/1310735
[19] Kohn M J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(46): 19691-19695. doi: 10.1073/pnas.1004933107
[20] Liu W G, Huang Y S. Reconstructing in-situ vegetation dynamics using carbon isotopic composition of biopolymeric residues in the central Chinese Loess Plateau [J]. Chemical Geology, 2008, 249(3-4): 348-356. doi: 10.1016/j.chemgeo.2008.01.010
[21] Liu W G, Yang H, Ning Y F et al. Contribution of inherent organic carbon to the bulk δ13C signal in loess deposits from the arid western Chinese Loess Plateau [J]. Organic Geochemistry, 2007, 38(9): 1571-1579. doi: 10.1016/j.orggeochem.2007.05.004
[22] 饶志国, 郭文康, 薛骞, 等. 黄土高原西部地区黄土地层有机质主要来源分析[J]. 第四纪研究, 2015, 35(4):819-827 doi: 10.11928/j.issn.1001-7410.2015.04.04 RAO Zhiguo, GUO Wenkang, XUE Qian et al. Assessment on primary provenance of organic matter in loess/paleosol sequences in the western Chinese Loess Plateau: local biomass or bedrocks in dust source regions? [J]. Quaternary Sciences, 2015, 35(4): 819-827. doi: 10.11928/j.issn.1001-7410.2015.04.04
[23] Zhang Z H, Zhao M X, Lu H Y et al. Lower temperature as the main cause of C4 plant declines during the glacial periods on the Chinese Loess Plateau[J]. Earth and Planetary Science Letters, 214(3-4): 467-481.
[24] 饶志国, 陈发虎, 曹洁, 等. 黄土高原西部地区末次冰期和全新世有机碳同位素变化与C3 /C4植被类型转换研究[J]. 第四纪研究, 2005, 25(1):107-114 doi: 10.3321/j.issn:1001-7410.2005.01.015 RAO Zhiguo, CHEN Fahu, CAO Jie et al. Variation of soil organic carbon isotope and C3/C4 vegetation type transition in the western loess plateau during the last glacial and Holocene periods [J]. Quaternary Sciences, 2005, 25(1): 107-114. doi: 10.3321/j.issn:1001-7410.2005.01.015
[25] Liu W G, Yang H, Sun Y B et al. δ13C values of loess total carbonate: A sensitive proxy for Asian summer monsoon in arid northwestern margin of the Chinese Loess Plateau [J]. Chemical Geology, 2011, 284(3-4): 317-322. doi: 10.1016/j.chemgeo.2011.03.011
[26] Yang S L, Ding Z L, Li Y Y et al. Warming-induced northwestward migration of the East Asian monsoon rain belt from the Last Glacial Maximum to the mid-Holocene [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(43): 13178-13183. doi: 10.1073/pnas.1504688112
[27] 林本海, 刘荣谟. 最近800ka黄土高原夏季风变迁的稳定同位素证据[J]. 科学通报, 1992(18):1691-1693 doi: 10.3321/j.issn:0023-074X.1992.18.023 LIN Benhai, LIU Rongmo. Stable isotopic evidence of the summer monsoon evolution during the last 800ka in Chinese Loess Plateau [J]. Chinese Science Bulletin, 1992(18): 1691-1693. doi: 10.3321/j.issn:0023-074X.1992.18.023
[28] Vidic N J, Montañez I P. Climatically driven glacial-interglacial variations in C3 and C4 plant proportions on the Chinese Loess Plateau [J]. Geology, 2004, 32(4): 337-340. doi: 10.1130/G20222.2
[29] Chen F H, Rao Z G, Zhang J W et al. Variations of organic carbon isotopic composition and its environmental significance during the last glacial on western Chinese Loess Plateau [J]. Chinese Science Bulletin, 2006, 51(13): 1593-1602. doi: 10.1007/s11434-006-2003-6
[30] Liu W G, Huang Y S, An Z S et al. Summer monsoon intensity controls C4/C3 plant abundance during the last 35 ka in the Chinese Loess Plateau: Carbon isotope evidence from bulk organic matter and individual leaf waxes [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 220(3-4): 243-254. doi: 10.1016/j.palaeo.2005.01.001
[31] Hatté C, Fontugne M, Rousseau D D et al. δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian [J]. Geology, 1998, 26(7): 583-586. doi: 10.1130/0091-7613(1998)026<0583:CVOLOM>2.3.CO;2
[32] 郑祥民, 刘飞. 长江三角洲与东海岛屿黄土研究综述[J]. 华东师范大学学报: 自然科学版, 2006(6):9-24 ZHENG Xiangmin, LIU Fei. Review of research on loess in the Yangtze River delta and the East China Sea islands [J]. Journal of East China Normal University(Natural Science), 2006(6): 9-24.
[33] 石娇星. 舟山群岛植被分类与制图[D]. 华东师范大学硕士学位论文, 2021: 62-63 SHI Jiaoxing. Vegetation classification and mapping of Zhoushan archipelago[D]. Master Dissertation of East China Normal University, 2021: 62-63.
[34] Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503-537. doi: 10.1146/annurev.pp.40.060189.002443
[35] 饶志国, 朱照宇, 贾国东, 等. 环北太平洋地区现代植被中C3/C4 植物相对丰度与气候条件关系研究[J]. 科学通报, 2010, 55(18):1931-1936 doi: 10.1007/s11434-010-3101-z RAO Zhiguo, ZHU Zhaoyu, JIA Guodong et al. Relationship between climatic conditions and the relative abundance of modern C3 and C4 plants in three regions around the North Pacific [J]. Chinese Science Bulletin, 2010, 55(18): 1931-1936. doi: 10.1007/s11434-010-3101-z
[36] Tieszen L L, Reed B C, Bliss N B et al. NDVI, C3 and C4 production and distributions in Great Plains grassland land cover classes [J]. Ecological Applications, 1997, 7(1): 59-78.
[37] Bird M I, Pousai P. Variations of δ 13C in the surface soil organic carbon pool [J]. Global Biogeochemical Cycles, 1997, 11(3): 313-322. doi: 10.1029/97GB01197
[38] 饶志国, 贾国东, 朱照宇, 等. 中国东部表土总有机质碳同位素和长链正构烷烃碳同位素对比研究及其意义[J]. 科学通报, 2008, 53(24):3921-3927 doi: 10.3321/j.issn:0023-074X.2008.17.013 RAO Zhiguo JIA Guodong ZHU Zhaoyu et al. Comparison of the carbon isotope composition of total organic carbon and long-chain n-alkanes from surface soils in Eastern China and their significance [J]. Chinese Science Bulletin, 2008, 53(24): 3921-3927. doi: 10.3321/j.issn:0023-074X.2008.17.013
[39] 何勇, 秦大河, 任贾文, 等. 塬堡黄土剖面末次间冰期古土壤有机质碳同位素记录的夏季风演化历史[J]. 科学通报, 2002, 47(15):1289-1291 doi: 10.3321/j.issn:0023-074X.2002.12.013 HE Yong, QIN Dahe, REN Jiawen et al. The summer monsoon evolution recorded by carbon isotope of organic matter from the Yuanbao loess section during the last Interglaciation [J]. Chinese Science Bulletin, 2002, 47(15): 1289-1291. doi: 10.3321/j.issn:0023-074X.2002.12.013
[40] Wang G A, Feng X, Han J et al. Paleovegetation reconstruction using δ13C of soil organic matter [J]. Biogeosciences, 2008, 5: 1325-1337. doi: 10.5194/bg-5-1325-2008
[41] Quade J, Cerling T E, Bowman J R. Development of Asian monsoon revealed by marked ecological shift during the latest Miocene in northern Pakistan [J]. Nature, 1989, 342(6246): 163-166. doi: 10.1038/342163a0
[42] Cerling T E. The stable isotopic composition of modern soil carbonate and its relationship to climate [J]. Earth and Planetary Science letters, 1984, 71(2): 229-240. doi: 10.1016/0012-821X(84)90089-X
[43] 刘卫国, 宁有丰, 安芷生, 等. 黄土高原现代土壤和古土壤有机碳同位素对植被的响应[J]. 中国科学D辑, 2005, 48(10):93-99 LIU Weiguo, NING Youfeng, AN Zhisheng, et al. Carbon isotopic composition of modern soil and paleosol as a response to vegetation change on the Chinese Loess Plateau [J]. Science in China Series D:Earth Sciences, 2005, 48(10): 93-99.
[44] Lyu A Q, Lu H Y, Zeng L et al. Vegetation variation of loess deposits in the southeastern Inner Mongolia, NE China over the past ~1.08 million years [J]. Journal of Asian Earth Sciences, 2018, 155: 174-179. doi: 10.1016/j.jseaes.2017.11.013
[45] 张月馨, 迟云平, 谢远云, 等. 中更新世以来哈尔滨黄土有机碳同位素组成及其古气候意义[J]. 地球学报, 2020, 41(4):525-534 doi: 10.3975/cagsb.2020.040602 ZHANG Yuexin, CHI Yunping, XIE Yuanyun et al. Organic carbon isotope composition of Harbin loess since the Mid-Pleistocene and its paleoclimatic significance [J]. Acta Geoscientica Sinica, 2020, 41(4): 525-534. doi: 10.3975/cagsb.2020.040602
[46] 匡欢传, 周浩达, 胡建芳, 等. 末次盛冰期和全新世大暖期湖光岩玛珥湖沉积记录的正构烷烃和单体稳定碳同位素分布特征及其古植被意义[J]. 第四纪研究, 2013, 33(6):1222-1233 doi: 10.3969/j.issn.1001-7410.2013.06.18 KUANG Huanchuan ZHOU Haoda, HU Jianfang et al. Variations of n-alkanes and compound-specific carbon isotopes in sediment from Huguangyan Maar Lake during the last glacial maximum and Holocence optimum: Implications for paleovegetation [J]. Quaternary Sciences, 2013, 33(6): 1222-1233. doi: 10.3969/j.issn.1001-7410.2013.06.18
[47] 顾兆炎, 刘强, 许冰, 等. 气候变化对黄土高原末次盛冰期以来的C3/C4 植物相对丰度的控制[J]. 科学通报, 2003, 48(12):1271-1276 doi: 10.3321/j.issn:0023-074X.2003.12.008 GU Zhaoyan, LIU Qiang, XU Bing et al. Climate as the dominant control on C3 and C4 plant abundance in the Loess Plateau: Organic carbon isotope evidence from the last glacial-interglacial loess-soil sequences [J]. Chinese Science Bulletin, 2003, 48(12): 1271-1276. doi: 10.3321/j.issn:0023-074X.2003.12.008
[48] Diefendorf A F, Mueller K E, Wing S L et al. Global patterns in leaf 13C discrimination and implications for studies of past and future climate [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(13): 5738-5743. doi: 10.1073/pnas.0910513107
[49] Ehleringer J R, Cooper T A. Correlations between carbon isotope ratio and microhabitat in desert plants [J]. Oecologia, 1988, 76(4): 562-566. doi: 10.1007/BF00397870
[50] 王国安, 韩家懋, 刘东生. 中国北方黄土区C-3草本植物碳同位素组成研究[J]. 中国科学D辑, 2003, 46(10):1069-1076 doi: 10.3321/j.issn:1006-9267.2003.06.008 WANG Guoan, HAN Jiamao, LIU Tungsheng. The carbon isotope composition of C3 herbaceous plants in loess area of northern China [J]. Science in China series D:Earth Sciences, 2003, 46(10): 1069-1076. doi: 10.3321/j.issn:1006-9267.2003.06.008
[51] Liu W G, Feng X H, Ning Y F et al. δ13C variation of C3 and C4 plants across an Asian monsoon rainfall gradient in arid northwestern China [J]. Global Change Biology, 2005, 11(7): 1094-1100. doi: 10.1111/j.1365-2486.2005.00969.x
[52] Wang G A, Li J Z, Liu X Z et al. Variations in carbon isotope ratios of plants across a temperature gradient along the 400 mm isoline of mean annual precipitation in north China and their relevance to paleovegetation reconstruction [J]. Quaternary Science Reviews, 2013, 63: 83-90. doi: 10.1016/j.quascirev.2012.12.004
[53] An Z S, Kukla G J, Porter S C et al. Magnetic susceptibility evidence of monsoon variation on the Loess Plateau of central China during the last 130, 000 years [J]. Quaternary Research, 1991, 36(1): 29-36. doi: 10.1016/0033-5894(91)90015-W
[54] 宁有丰, 刘卫国, 安芷生. 甘肃西峰黄土-古土壤剖面的碳酸盐与有机碳的碳同位素差值(Δδ13C)的变化及其古环境意义[J]. 科学通报, 2006, 51(11):1350-1354 doi: 10.1007/s11434-006-1350-7 NING Youfeng, LIU Weiguo, AN Zhisheng. Variation of soil Δδ13C values in Xifeng loess-paleosol sequence and its paleoenvironmental implication [J]. Chinese Science Bulletin, 2006, 51(11): 1350-1354. doi: 10.1007/s11434-006-1350-7
[55] Wang G A, Zhang L L, Zhang X Y et al. Chemical and carbon isotopic dynamics of grass organic matter during litter decompositions: A litterbag experiment [J]. Organic Geochemistry, 2014, 69: 106-113. doi: 10.1016/j.orggeochem.2014.02.012
[56] Feng X H, Epstein S. Carbon isotopes of trees from arid environments and implications for reconstructing atmospheric CO2 concentration [J]. Geochimica et Cosmochimica Acta, 1995, 59(12): 2599-2608. doi: 10.1016/0016-7037(95)00152-2
[57] Jouzel J, Lorius C, Petit J R et al. Vostok ice core: a continuous isotope temperature record over the last climatic cycle(160 000 years) [J]. Nature, 1987, 329(6138): 403-408. doi: 10.1038/329403a0
[58] SeltzerA M, Ng J, Aeschbach W et al. Widespread six degrees Celsius cooling on land during the Last Glacial Maximum [J]. Nature, 2021, 593(7858): 228-232. doi: 10.1038/s41586-021-03467-6
[59] 陈骏, 汪永进, 季峻峰, 等. 陕西洛川黄土剖面的Rb/Sr值及其气候地层学意义[J]. 第四纪研究, 1999, 19(4):350-356 doi: 10.3321/j.issn:1001-7410.1999.04.007 CHEN Jun, WANG Yongjin, JI Junfeng et al. Rb/Sr variations and its climatic stratigraphical significance of a loess-paleosol profile from Louchuan, Shanxi province [J]. Quaternary Sciences, 1999, 19(4): 350-356. doi: 10.3321/j.issn:1001-7410.1999.04.007
[60] Peng W B, Nie J S, Wang Z et al. A major change in precipitation gradient on the Chinese Loess Plateau at the Pliocene-Quaternary boundary [J]. Journal of Asian Earth Sciences, 2018, 155: 134-138. doi: 10.1016/j.jseaes.2017.10.031
[61] Hatté C, Antoine P, Fontugne M et al. δ13C of loess organic matter as a potential proxy for paleoprecipitation [J]. Quaternary Research, 2001, 55(1): 33-38. doi: 10.1006/qres.2000.2191
[62] Wang Y J, Cheng H, Edwards R L et al. A high-resolution absolute-dated late Pleistocene monsoon record from Hulu Cave, China [J]. Science, 2001, 294(5550): 2345-2348. doi: 10.1126/science.1064618
[63] Linsley B K. Oxygen-isotope record of sea level and climate variations in the Sulu Sea over the past 150, 000 years [J]. Nature, 1996, 380(6571): 234-237. doi: 10.1038/380234a0
[64] Petit J R, Jouzel J, Raynaud D et al. Climate and atmospheric history of the past 420, 000 years from the Vostok ice core, Antarctica [J]. Nature, 1999, 399(6735): 429-436. doi: 10.1038/20859
[65] 饶志国, 陈发虎, 张晓, 等. 末次冰期以来全球陆地植被中C3/C4植物相对丰度时空变化基本特征及其可能的驱动机制[J]. 科学通报, 2012, 57(31):4024-4035 doi: 10.1007/s11434-012-5233-9 RAO Zhiguo, CHEN Fahu, ZHANG Xiao et al. Spatial and temporal variations of C3/C4 relative abundance in global terrestrial ecosystem since the Last Glacial and its possible driving mechanisms [J]. Chinese Science Bulletin, 2012, 57(31): 4024-4035. doi: 10.1007/s11434-012-5233-9
-
期刊类型引用(1)
1. 王丹,RASHID Harunur. 长链烯酮在西北大西洋重建全新世气候变化的研究进展. 海洋科学. 2022(01): 181-191 .  百度学术
百度学术
其他类型引用(3)
计量
- 文章访问数: 1104
- HTML全文浏览量: 368
- PDF下载量: 142
- 被引次数: 4


