
Spatial-temporal variation and salt transport in the intermediate water of the northern South China Sea
-
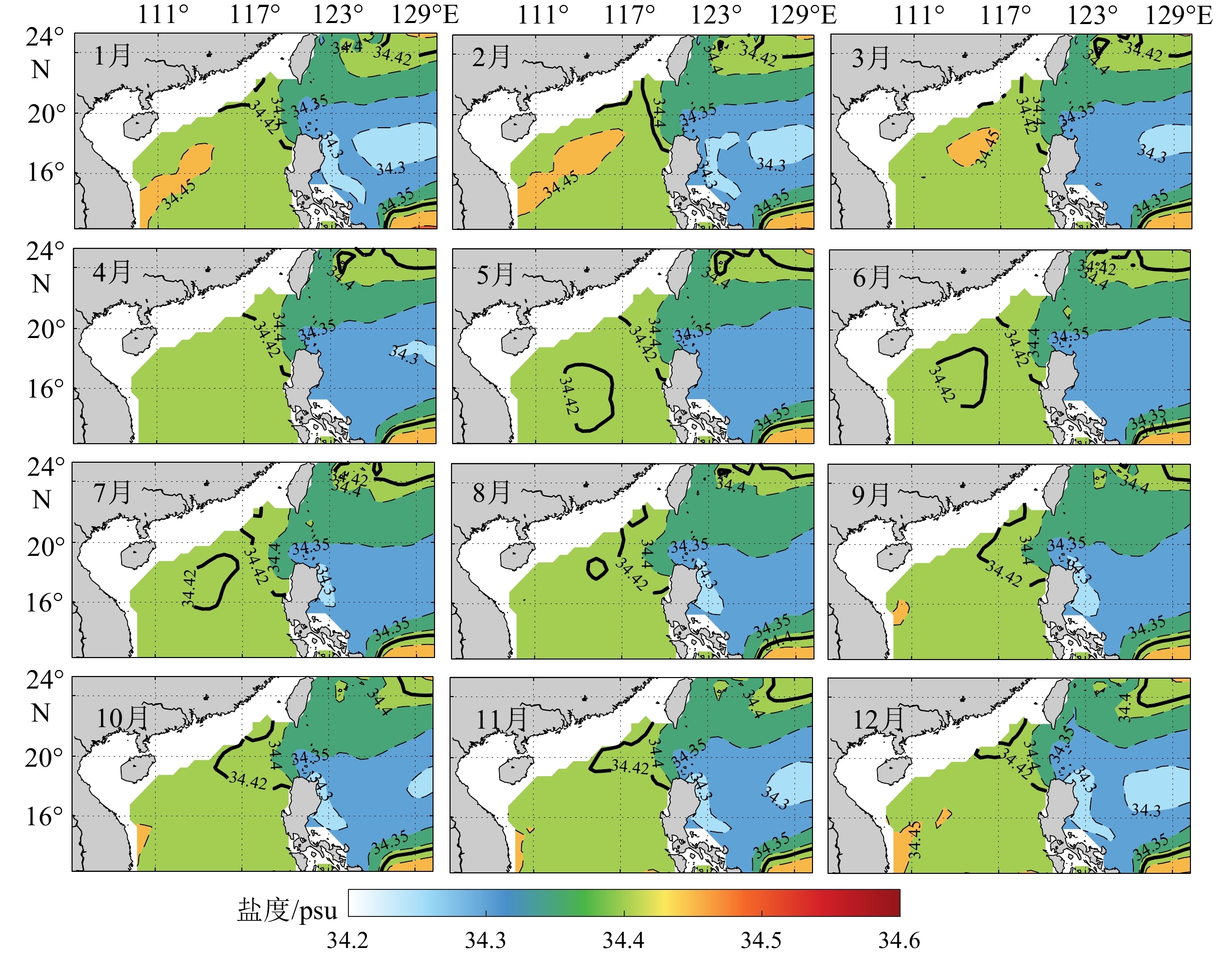
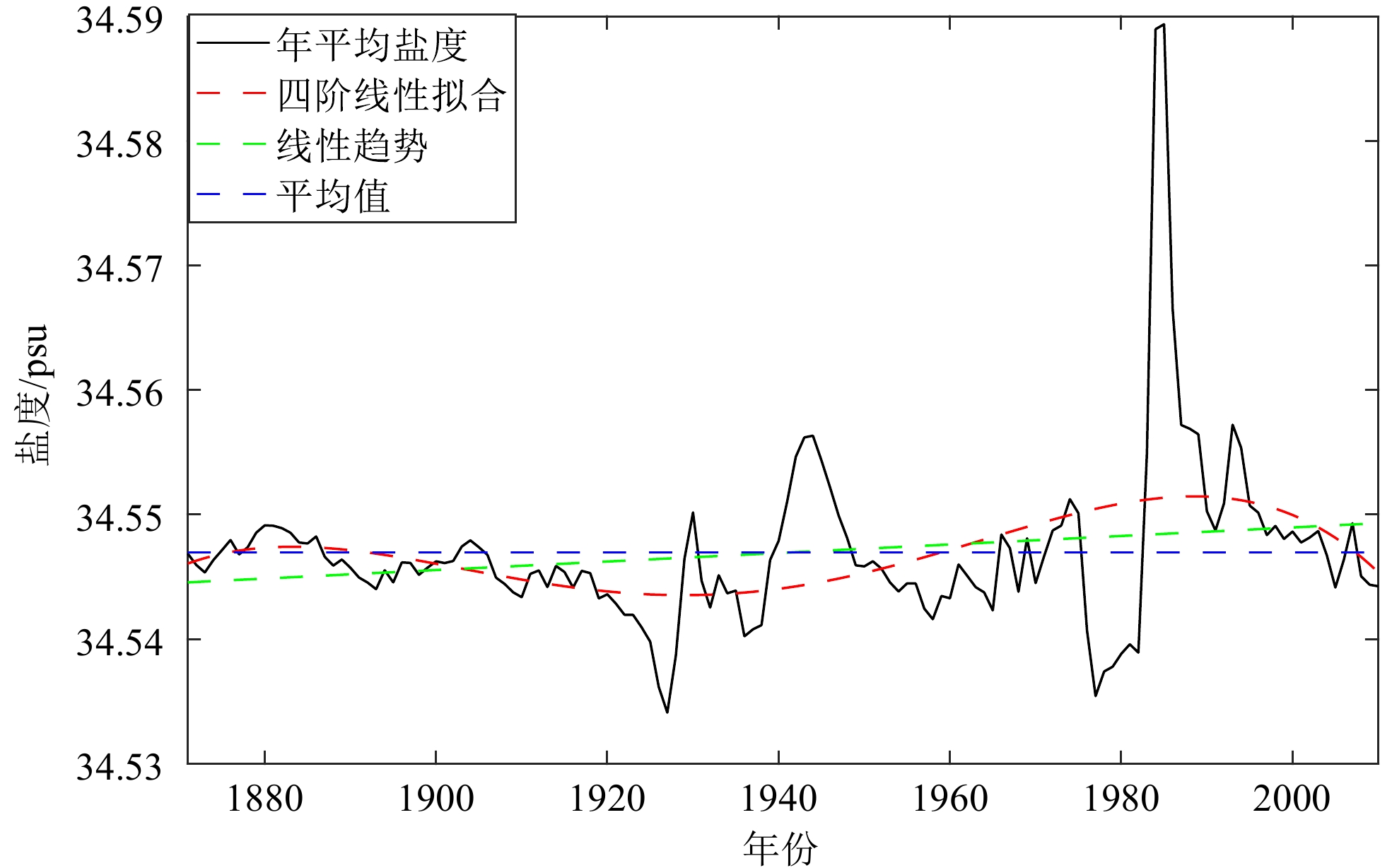
摘要: 为探讨南海北部中层水盐度的时空分布特征及与外海水的交换,选用1871—2010年共140年的月平均海洋同化数据(SODA2.2.4),利用EOF、小波等分析方法,分析南海北部中层水盐度的时空分布、变化周期、盐通量等特征。结果表明:① 南海北部中层水的盐度月际变化特征为:1—3月,南海北部中层水受到越南东部沿岸向东北方向延伸出的高盐舌的影响(>34.45 psu),盐度偏高,中部盐度较低;4月南海北部中层水的盐度分布均一,盐度范围在34.40~34.45 psu;5—8月,夏季南海北部的中层气旋式环流将北太平洋中层水(North Pacific Intermediate Water,NPIW)携带至南海中部,在南海中部偏北形成一低盐水团(<34.42 psu), 并在此时间段内低盐水团逐渐向东北方向移动,直至9—10月,上述低盐水团与NPIW混合;11—12月,NPIW东撤,收缩至吕宋海峡西北侧。② 过去140年,南海北部中层水的盐度值波动上升,其中,1985年中层水盐度值最高,可能与当年夏季风爆发早且强有关。1963—2002年,存在较强的16~21 a的周期性,小波系数的实部正相位的峰值和负相位的谷值所对应的年份,与ENSO冷事件具有较好的对应。③ 对EOF第一模态分析显示,整个研究海域为单极子变化特征,呈一致性变化。第二模态的空间分布呈偶极子形态,一正一负相位中心沿NW-SE走向对称分布。④ 南海北部中层水全年的净盐通量均由南海向东流入太平洋,夏季最少,冬季最多。1871—2010年的140年间,中层水的盐通量一直处于下降的趋势,下降的趋势十分缓慢。Abstract: To explore the spatial-temporal distribution of salinity in the intermediate water and water exchange with the external waters of the northern South China Sea(SCS), the monthly mean oceanic assimilation data of past 140-year from 1871 to 2010 (SODA2.2.4) were selected, on which the spatial-temporal variation, periodicity, and salt transport of intermediate water salinity in the SCS and adjacent northwest Pacific Ocean were analyzed using wavelet analysis and EOF decomposition. The result show that the monthly salinity variation of intermediate water in the northern SCS is higher in January-March than that of the central part due to northeastward extending of high salinity water from the eastern coast of Vietnam (>34.45 psu). The salinity distribution of the intermediate water in the northern SCS is uniform in April, and the salinity ranged from 34.40 to 34.45 psu. In May-August, there is a cyclonic circulation in the intermediate water of the northern SCS in summer, which can carry the North Pacific Intermediate Water (NPIW) to the central part of SCS, and then forms a low-salinity water lens (<34.42 psu) in the north-central SCS. Meanwhile, the low-salinity water lens move northeastward gradually until mix with NPIW in September-October. In November-December, NPIW withdraws eastward and shrinks to the northwest of the Luzon Strait. In the past 140 years, the salinity of intermediate water in the northern SCS has been rising periodically, reaching the peak in 1985, which may be related to the early and strong summer monsoon. From 1963 to 2002, a strong 16-21–year periodicity is shown, and the years of the maximum (minimum) values of the real part of wavelet coefficients well agree with corresponding ENSO cold events. The first EOF analysis shows that the intermediate water salinity in the northern SCS is in a monopole and consistent variation pattern. The positive and negative phase centers identified by the second mode is symmetrical in distribution in NW-SE strike. The annual net salt fluxes of the intermediate water in the northern SCS flow eastward from the South China Sea to the Pacific Ocean, the least in summer and the most in winter. During the 140 years from 1871 to 2010, the salt fluxes in the intermediate water has been decreasing constantly and slowly.
-
Keywords:
- intermediate water /
- Northwest Pacific /
- seawater salinity /
- salt transport /
- the South China Sea
-
北冰洋被高耸的洋中脊—罗蒙诺索夫脊分隔为东、西两部分。现代北冰洋的显著特点之一是海水的分层结构[1],它拥有以下四大主要水团:极地表层水(PSW,水深0~50 m,水温−2~0 °C,盐度32~34 psu),大西洋水(AW,水深200~1000 m,水温≥0 °C,盐度34.3~34.8 psu),北冰洋中层水(AIW,水温−0.5~0 °C,盐度34.6~34.8 psu),以及北冰洋深层水(AODW);AODW 分为加拿大海盆深层水(CBDW,水深>1500 m,水温−0.5~−0.3 °C,盐度34.95 psu)和欧亚海盆深层水(EBDW,水深>2 000 m,水温−1.0~−0.6 °C,盐度34.94 psu)[2-5]。AW和PSW被一层发育良好的盐跃层(水深<200 m)所分隔[6]。温度和盐度较高的北大西洋水体经由弗拉姆海峡和巴伦支海流入北冰洋,潜入相对低盐的盐跃层之下并逆时针流动[4,7],形成了位于中层的AW和AIW[8],因此北大西洋是北冰洋重要的热量和盐分供给源。在北冰洋,大尺度的表层洋流有从西伯利亚陆架流向弗拉姆海峡的穿极洋流,以及加拿大海盆的波弗特环流(图1a)。北冰洋从欧亚大陆和北美大陆接受了约占全球总量10%的淡水流量[9],而波弗特环流夹裹了大量的淡水[10-12]。阿尔法脊的表层水被常年海冰所覆盖[13],同时也处于波弗特环流的影响之下。
![]() 图 1 西北冰洋阿尔法脊B84A岩芯平面位置(a)以及在海洋水团中的位置(b)淡黄色圆点表示本文提及的其他3个岩芯站位(详细信息见表1)。蓝色和粉红色点线分别表示1979—2006年间的平均海冰范围及近年(2012年数据)最小海冰范围[22],浅白色阴影部分表示推定的更新世冰盖范围[23-24]。EAIS-欧亚冰盖,GIS-格陵兰冰盖,LIS-劳伦太德冰盖,ESIS-东西伯利亚冰盖,EI-埃尔斯米尔岛,CAA-加拿大北极群岛,CB-加拿大海盆,EB-欧亚海盆,MB-马卡罗夫海盆,AR-阿尔法脊,LR-罗蒙诺索夫脊,CP-楚科奇海台,NR-北风脊,BG-波弗特环流,TPD-穿极洋流,PSW-极地表层水,AW-大西洋水,AIW-北冰洋中层水,AODW-北冰洋深层水。Figure 1. Location of core B84A in a map view (a) and cross section (b)Section runs from the Eurasian Basin at point A, across to the Canada Basin at point B. The location where the core B84A was drilled is marked by red circle with white outline, and other cores mentioned in this paper are represented by light yellow circles (see Table 1 for more core information). Blue and pink dashed lines indicate climatological average (1979 to 2006) and recent minimum (2012) September sea ice extent, respectively[22]. Light shaded areas indicate tentative extent of Pleistocene glaciations around the Arctic Ocean[23-24]. Abbreviations: EAIS (Eurasian Ice Sheet), GIS (Greenland Ice Sheet), LIS (Laurentide Ice Sheet), ESIS (East Siberian Ice Sheet), EI (Ellesmere Island), CAA (Canadian Arctic Archipelago), CB (Canada Basin), EB (Eurasia Basin), MB (Makarov Basin), AR (Alpha Ridge), LR (Lomonosov Ridge), CP (Chukchi Plateau), NR (Northwind Ridge), BG (Beaufort Gyre), TPD (Transpolar Drift), PSW (Polar Surface Water), AW (Atlantic Water), AIW (Arctic Intermediate Water), and AODW (Arctic Ocean Deep Water).
图 1 西北冰洋阿尔法脊B84A岩芯平面位置(a)以及在海洋水团中的位置(b)淡黄色圆点表示本文提及的其他3个岩芯站位(详细信息见表1)。蓝色和粉红色点线分别表示1979—2006年间的平均海冰范围及近年(2012年数据)最小海冰范围[22],浅白色阴影部分表示推定的更新世冰盖范围[23-24]。EAIS-欧亚冰盖,GIS-格陵兰冰盖,LIS-劳伦太德冰盖,ESIS-东西伯利亚冰盖,EI-埃尔斯米尔岛,CAA-加拿大北极群岛,CB-加拿大海盆,EB-欧亚海盆,MB-马卡罗夫海盆,AR-阿尔法脊,LR-罗蒙诺索夫脊,CP-楚科奇海台,NR-北风脊,BG-波弗特环流,TPD-穿极洋流,PSW-极地表层水,AW-大西洋水,AIW-北冰洋中层水,AODW-北冰洋深层水。Figure 1. Location of core B84A in a map view (a) and cross section (b)Section runs from the Eurasian Basin at point A, across to the Canada Basin at point B. The location where the core B84A was drilled is marked by red circle with white outline, and other cores mentioned in this paper are represented by light yellow circles (see Table 1 for more core information). Blue and pink dashed lines indicate climatological average (1979 to 2006) and recent minimum (2012) September sea ice extent, respectively[22]. Light shaded areas indicate tentative extent of Pleistocene glaciations around the Arctic Ocean[23-24]. Abbreviations: EAIS (Eurasian Ice Sheet), GIS (Greenland Ice Sheet), LIS (Laurentide Ice Sheet), ESIS (East Siberian Ice Sheet), EI (Ellesmere Island), CAA (Canadian Arctic Archipelago), CB (Canada Basin), EB (Eurasia Basin), MB (Makarov Basin), AR (Alpha Ridge), LR (Lomonosov Ridge), CP (Chukchi Plateau), NR (Northwind Ridge), BG (Beaufort Gyre), TPD (Transpolar Drift), PSW (Polar Surface Water), AW (Atlantic Water), AIW (Arctic Intermediate Water), and AODW (Arctic Ocean Deep Water).Haley等[14]对北冰洋岩芯中的钕同位素分析结果显示,北冰洋海水的分层格局形成于早更新世,但中层水体(包括AW和AIW)的性质随气候而变化:间冰期,中层水体主要来自北大西洋中层水的贡献,而冰期则主要来自于在欧亚大陆架上形成的卤水。另一方面,Cronin等[15]利用介形虫壳体 Mg/Ca古温度计重建了50 ka以来北冰洋的水团变化历史,得出了末次冰期中盐跃层增厚,AW在整个北冰洋均向下被挤压到1000~2500 m深度的著名结论。然而,我们发现Cronin等的结论虽然可以很好地解释末次冰期中介形虫化石群在北冰洋罗蒙诺索夫脊以东的分布方式,却无法解释它们在罗蒙诺索夫脊以西的分布。
介形虫是北冰洋微体化石的重要门类之一。Gemery等[16]基于整个北冰洋1000多个表层沉积样数据而建立的北冰洋介形虫数据库(Arctic Ostracoda Database, AOD)表明,介形虫属种与海洋环境之间存在着明确的对应关系。自20世纪80年代以来,介形虫在北冰洋新近纪—第四纪古海洋学中的应用越来越多,揭示了介形虫化石群与古气候事件的关联性[17]。在过去几年里,本文作者对中国北极科学考察期间“雪龙号”科考船在西北冰洋楚科奇海台一带钻取的3根岩芯(水深<1000 m)中的介形虫及部分底栖有孔虫属种进行详细研究,并对前人的北冰洋介形虫化石数据[18-19]进行重新分析后,得出了如下结论:在包括末次冰期在内的 MIS 5–2期,AW在西北冰洋并非迁移到更深水域,而是因受罗蒙诺索夫脊的阻隔而变弱甚或消失了[17,20]。
我们的上述结论是对Cronin等[15]结论的重要修正,揭示了气候寒冷期里东、西北冰洋迥异的水体结构,它有待更多的研究来进一步证实。这同时也衍生出了新的科学问题:在更深水域中,北冰洋的水团是否也随气候冷暖交替而发生变化?作为解答这一问题的第一步,本研究选取了中国第三次北极科学考察期间在北冰洋中部阿尔法脊采取的岩心ARC3-B84A(站位水深2280 m),对其中的介形虫化石群进行了详细分析。研究内容包含两个方面:首先,基于北冰洋介形虫数据库(AOD)[16],对ARC3-B84A岩芯中介形虫主要属种在现代北冰洋的生态分布,尤其与水团的对应关系进行了分析;然后,根据主要属种在该岩芯中的时代分布,重建了MIS 13期以来深部水团在阿尔法脊一带海域的变迁历史。
1. 材料及方法
1.1 研究材料
本研究材料来自中国第三次北极科学考察[21]于2008年夏季在阿尔法脊采集的ARC3-B84A岩芯(以下简称B84A岩心)。该岩芯长188 cm,站位水深2280 m,位于AODW上部;与之相比较,用于西北冰洋中层水团研究的3根岩芯(ARC3-P31、ARC6-R14及ARC7-P12)[17]均采自楚科奇海台,站位处于现代北冰洋AW之内(图1,表1)。
表 1 本研究使用的岩芯及其信息汇总Table 1. Information for all the cores used in this study1.2 研究方法
本研究采用标准微体古生物学分析方法对B84A岩芯沉积物样品进行处理。岩芯按2 cm间隔取样,共获得94个样品。在同济大学海洋地质国家重点实验室,将分割的样品用烘箱烘干、称重,然后经水泡分散后,用孔径63 μm的铜筛水洗。将>63 μm的屑样烘干后,再用孔径150 μm的筛子干筛。在体视显微镜下,将>150 μm屑样中的介形虫个体全部挑出用于属种鉴定统计分析。本文主要采用Yasuhara等[25]、Gemery等[16]作为北冰洋介形虫分类鉴定标准。此外,利用同济大学海洋地质国家重点实验室的扫描电子显微镜(TESCAN Mira 3)对介形虫标本进行观察并拍摄照片。
本研究计算了每个样品中的介形虫丰度(每克干样中介形虫的壳瓣数),因为它反映了底栖生物古生产力变化;计算了常见属种在每个样品中的个体数量百分比,以便复原古水团变迁。介形虫个体数用单瓣数来表示,1个合瓣计为2个单瓣。在计算各属种的百分比时,为了减少误差,本文将个体数量较少的相邻样品合并起来,以使合并后的瓣数大于50。
为了准确把握介形虫属种的水深分布及其与水团的对应关系,本文使用软件Ocean Data View (ODV)[26]分析了B84A岩芯中的常见种在现代北冰洋的分布。共计选取了北冰洋638个表层沉积物样品的介形虫数据,并将数据输入ODV软件绘制各个属种在现代北冰洋的丰度等值线断面图。这些样品由AOD[16]的1340个样品中的626个,以及我们自己拥有的231个西北冰洋样品中的12个构成,每个样品都包含了50个以上介形虫个体。
2. 地层年代框架
B84A岩芯沉积物主要由粉砂质黏土构成。Wang等[27]综合运用AMS14C测年、层序对比、旋回地层学校准等方法,建立了该岩芯可靠的年代框架。具体步骤如下:首先基于AMS 14C测年、Ca和Mn元素峰值、地层标志种的分布等,确定氧同位素期次;然后把B84A岩芯的Ca和Mn的stack与LR04深海氧同位素曲线[28]对比,确定年龄控制点后转换为深度–年龄模式;最后分别通过“插值法”获得每个2 cm间隔岩芯样品的年龄。根据年龄模式,该岩芯包含了MIS 1期及MIS 3–13期的沉积记录,MIS 2期缺失。
3. 结果及讨论
3.1 B84A岩芯中的介形虫及其生态特性
阿尔法脊B84A岩芯中介形虫化石数量丰富,从94个样品中共挑出介形虫7011瓣,鉴定出8属11种。化石群的优势种为Polycope spp.(占总个体数量的52.2%)和Cytheropteron sedovi Schneider 1969(占20.7%),常见种为Cytheropteron scoresbyi Whatley and Coles 1987(占4.9%)、Acetabulastoma arcticum Schornikov 1970(占7.0%)、Henryhowella asperrima (Reuss 1850)(占5.5%)、Microcythere medistriatum Joy and Clark 1977(占1.8%)、Pseudocythere caudata Sars 1866(占1.9%)、Cytheropteron higashikawai Ishizaki1981(占1.8%)以及Pedicythere spp.(占1.0%)。有趣的是,北冰洋深海介形虫的代表属之一Krithe[17-19],在该岩芯中仅发现6瓣;而北冰洋大西洋水(AW)的指示者Cytheropteron perlaria Hao 1988[17],在该岩芯中仅出现1瓣(42~44 cm孔深处)。图2为部分属种的扫描电子显微镜照片。
![]() 图 2 阿尔法脊B84A岩芯中介形虫标本扫描电子显微镜照片a. Polycope biretculata Joy and Clark 1977,右壳瓣;b. Polycope inornata Joy and Clark 1977,右壳瓣;c. Polycope horida Joy and Clark 1977,右壳瓣;d. Polycope moenia Joy and Clark 1977,右壳瓣;e. Polycope arcys Joy and Clark 1977,左壳瓣;f. Acetabulastoma arctium Schornikov 1970,左壳瓣;g. Cytheropteron scoresbyi Whatley and Coles 1987,左壳瓣;h. Cytheropteron sedovi Schneider 1969,右壳瓣;i. Microythere medistriatum Joy and Clark 1977,左壳瓣;j. Pedicythere neofluitans Joy and Clark 1977,右壳瓣;k. Henryhowella asperrima (Reuss 1850),右壳瓣;l. Pseudocyhere caudata Sars 1866,右壳瓣。Figure 2. SEM photographs of ostracodes from core B84Aa. Polycope biretculata Joy and Clark 1977, RV; b. Polycope inornata Joy and Clark 1977, RV; c. Polycope horida Joy and Clark 1977, RV; d. Polycope moenia Joy and Clark 1977, RV; e. Polycope arcys Joy and Clark 1977, LV; f. Acetabulastoma arctium Schornikov 1970, LV; g. Cytheropteron scoresbyi Whatley and Coles 1987, LV; h. Cytheropteron sedovi Schneider 1969, RV; i. Microythere medistriatum Joy and Clark 1977, LV; j. Pedicythere neofluitans Joy and Clark 1977, RV; k. Henryhowella asperrima (Reuss 1850), RV; l. Pseudocyhere caudata Sars 1866, RV.
图 2 阿尔法脊B84A岩芯中介形虫标本扫描电子显微镜照片a. Polycope biretculata Joy and Clark 1977,右壳瓣;b. Polycope inornata Joy and Clark 1977,右壳瓣;c. Polycope horida Joy and Clark 1977,右壳瓣;d. Polycope moenia Joy and Clark 1977,右壳瓣;e. Polycope arcys Joy and Clark 1977,左壳瓣;f. Acetabulastoma arctium Schornikov 1970,左壳瓣;g. Cytheropteron scoresbyi Whatley and Coles 1987,左壳瓣;h. Cytheropteron sedovi Schneider 1969,右壳瓣;i. Microythere medistriatum Joy and Clark 1977,左壳瓣;j. Pedicythere neofluitans Joy and Clark 1977,右壳瓣;k. Henryhowella asperrima (Reuss 1850),右壳瓣;l. Pseudocyhere caudata Sars 1866,右壳瓣。Figure 2. SEM photographs of ostracodes from core B84Aa. Polycope biretculata Joy and Clark 1977, RV; b. Polycope inornata Joy and Clark 1977, RV; c. Polycope horida Joy and Clark 1977, RV; d. Polycope moenia Joy and Clark 1977, RV; e. Polycope arcys Joy and Clark 1977, LV; f. Acetabulastoma arctium Schornikov 1970, LV; g. Cytheropteron scoresbyi Whatley and Coles 1987, LV; h. Cytheropteron sedovi Schneider 1969, RV; i. Microythere medistriatum Joy and Clark 1977, LV; j. Pedicythere neofluitans Joy and Clark 1977, RV; k. Henryhowella asperrima (Reuss 1850), RV; l. Pseudocyhere caudata Sars 1866, RV.基于来自北冰洋638个表层沉积物样品的介形虫数据,本文分析了B84A中的2个优势种和7个常见种在现代北冰洋的分布,以便确定它们与海洋环境、尤其与水团的对应关系。9个属种的百分含量等值线图用ODV绘制,投影在连接加拿大海盆和巴伦支海的横断面上;同时还绘制了介形虫百分含量-水深散点图(图3),为介形虫属种与特定海洋环境的对应关系提供了新认知。9个属种与现代北冰洋水团的对应关系归纳如下(表2)。
表 2 B84A岩芯中的介形虫优势种和常见种在现代北冰洋各水团中的百分含量Table 2. Relative frequencies (%) of ostracode taxa, which are abundant or common in core B84A, in the Arctic water masses种名 0~50 m
(PSW)50~200 m
(盐跃层)200~1000 m
(AW)1000~2 000 m
(AIW)>2 000 m
(AODW)Polycope spp. 0.38 1.47 8.44 32.67 6.82 Microcythere medistriatum 0 0 0.20 2.07 1.92 Pseudocythere caudata 0.03 0.35 1.39 2.78 1.67 Pedicythere spp. 0 0.05 0.21 1.12 0.64 Henryhowella asperrima 0 0 0 1.42 2.90 Cytheropteron scoresbyi 0 0.39 3.64 8.05 10.47 Cytheropteron sedovi 0 0.02 1.11 9.72 19.16 Cytheropteron higashikawai 0 0.42 2.31 2.97 5.80 (1)北冰洋中层水(AIW)属种:Polycope spp.、Pseudocythere caudata和Pedicythere spp.的百分含量在1 000~2 000 m水深范围内(即AIW内)最高,但在该深度以上和以下明显降低(图3a, c, d;表2)。
![]() 图 3 B84A岩芯中介形虫主要属种在现代北冰洋的分布BS:巴伦支海,N-GR:南森-迦凯脊,LR:罗蒙诺索夫脊,MR:门捷列夫脊,CB:加拿大海盆。Figure 3. Distribution of key ostracode taxa in modern Arctic OceanAbbreviations: BS (Barents Sea), N-GR (Nansen-Gakkel Ridge), LR (Lomonosov Ridge), MR (Mendeleev Ridge), CB (Canada Basin).
图 3 B84A岩芯中介形虫主要属种在现代北冰洋的分布BS:巴伦支海,N-GR:南森-迦凯脊,LR:罗蒙诺索夫脊,MR:门捷列夫脊,CB:加拿大海盆。Figure 3. Distribution of key ostracode taxa in modern Arctic OceanAbbreviations: BS (Barents Sea), N-GR (Nansen-Gakkel Ridge), LR (Lomonosov Ridge), MR (Mendeleev Ridge), CB (Canada Basin).Polycope spp.营底栖生活,它们在北冰洋代表着一类机会种,在合适的环境中能够快速繁衍[29-30]。以往的研究已经发现Polycope与AIW之间存在着对应关系,并将其应用于古海洋学研究[17-19, 29, 31-32]。本研究对现代北冰洋数据的分析结果,再次证实了该属对AIW的适应性。
Pseudocythere caudata最早发现于北大西洋的潮下带[33],但是它在北冰洋栖息的水深范围达到3000 m,且主要分布在AIW中。以前的研究已经发现P. caudata与Polycope spp.在化石群中存在着共现关系[29, 34],并发现该种偏好AIW和AODW[17, 19]。
Pedicythere属在北冰洋有两个种,即P. neofluitans Joy and Clark 1977和P. arctica Yasuhara et al. 2014。该属零星分布于100~3400 m水深范围,百分含量通常小于2%,最大丰度出现于1000~2000 m水深范围,即AIW内。
(2)北冰洋深层水(AODW)属种:Cytheropteron sedovi、Henryhowella asperrima以及C. higashikawai的百分含量在>2 000 m水深范围(即AODW内)最高,而在其他水深范围内明显降低(图3h, e, i;表2)。在AODW内,C. sedovi和C. higashikawai主要分布于>3000 m水深范围,显示它们适应AODW下部;而H. asperrima则主要分布于2 000~3000 m范围,说明该种适应AODW上部环境。
(3)北冰洋中层水和深层水(AIW & AODW)属种:在>1000 m水深范围(即AIW和AODW内),Microcythere medistriatum和Cytheropteron scoresbyi的百分含量一直保持较高值,而在<1000 m水深则明显降低(图3b, g;表2)。本研究的分析结果与之前的研究结论[17]一致,即M. medistriatum和C. scoresbyi偏好AIW和AODW。
Acetabulastoma arcticum的现生分布仅限于北冰洋中央水域,其百分含量不超过10%(图3f)。该种是一个特例,因为它并非底栖种,而是附着在端足类Gammarus wilkitzkii体表的寄生种,这种端足类在整个生活史都处于北冰洋海冰卤水通道(brine channels)及融化穴(melting holes)中。因此,A. arcticum被视作北冰洋及周边海域常年海冰的指示种[16, 30, 35],它的分布与水团无关。
以上结果表明,B84A岩芯中的9个介形虫优势属种和常见属种中,除A. arcticum之外,其余8个属种的现生分布均与北冰洋特定的水团相关联,因此可以用于古水团重建的研究。
3.2 MIS 13期以来介形虫化石群变化特征及其影响因素
3.2.1 古生产力与海冰变化
B84A岩芯的94个样品中,介形虫平均丰度为2.9瓣/g,但丰度在不同时期有明显差异。介形虫丰度变化基本上与浮游、底栖有孔虫的丰度变化趋势同步(图4):总体上,丰度在MIS 13–10期很低,平均值仅为0.4瓣/g,而在MIS 9期之后显著升高,平均达到4.3瓣/g。MIS 10期之后,介形虫丰度的高值出现在间冰期的MIS 9、7、5、3、1期,低值则出现在冰期的MIS 8、6、4、2期。微体化石在间冰期多、冰期少的变化趋势在北冰洋似乎是普遍规律[17, 30, 36-38]。影响北冰洋沉积物中方解石质微体化石丰度的因素有若干个,包括沉积速率、溶蚀作用、生物生产力等;这些因素中,生物生产力似乎是最重要因素,因为本研究(图4)和以往的研究[30]都说明介形虫丰度与氧同位素曲线之间存在着非常好的相关性。本研究结果也证明了北冰洋> 2000 m水域底栖动物的生产力与较浅水域一样,受冰期–间冰期冷暖变化的影响。
![]()
在MIS 13–10期,介形虫和底栖、浮游有孔虫丰度均呈现显著的低值现象。在年代较老的地层中,溶蚀作用可能会损耗更多的介形虫壳体,造成丰度降低的假象,但是我们认为在B84A中并不存在这种可能性,理由有两个:首先,介形虫壳体保存状态在MIS 9期之前和之后没有可辨识的差异,而且壳体相对较薄且易碎的Polycope spp.在MIS 12–11期也有较高的百分含量(图4),说明介形虫化石群在这一时期的保存条件一直是良好的;第二,沉积速率在MIS 13–10期(平均0.42 cm/ka)和MIS 9–1期(平均0.37 cm/ka)之间没有明显差别。因此,有孔虫和介形虫在MIS13–10期的低丰度现象,可能是该时期海洋古生产力显著降低的真实反映。当然,这一推测尚有待北冰洋中部晚第四纪总有机碳分析数据的佐证。
Acetabulastoma arcticum是北冰洋常年海冰的指示种。图4显示A. arcticum在MIS 13–10期几近缺失,在孔深116 cm处(对应MIS 9期)开始较多地出现,因此MIS 9期很可能是常年海冰在罗蒙诺索夫脊以西最早出现的时期,正如Cronin等[30]所指出的那样。然而,尚难以解释为何在常年海冰不存在的MIS 13–10期,由有孔虫和介形虫所代表的海洋古生产力反而降低了。MIS 9之后,A. arcticum在冰期里(MIS 8、6、4期)非常稀少甚至缺失,但在间冰期(MIS 9、7、5、3期)通常占了介形虫总个体数的5%以上,最高达到>30%。在北冰洋中部[18]和西部[30]岩芯记录中,我们发现A. arcticum具有与B84A岩芯中相同的分布方式,即间冰期出现、冰期消失,说明该种及其寄主只能生活在常年海冰和季节性海冰共存的间冰期,而不能生活在洋面完全冻结的冰期环境。
3.2.2 各属种百分含量与古水团变迁
B84A岩芯中,占优势的介形虫属种是Polycope spp.(AIW指示种)和Cytheropteron sedovi(AODW指示种)。MIS 13期以来,这两个属种的百分含量曲线几乎呈镜像对称,二者呈负相关关系且发生了4次较大的转折,据此可以将岩芯记录分为5段,即MIS 13–12期、MIS 11–10期、MIS 9–MIS 5早期、MIS 5中–晚期以及MIS 4–1期。伴随着以上两个优势属种的此消彼长,其他6个与水团相关联的常见种(C. scoresbyi, C. higashikawai, M. medistriatum, Pseudocythere caudata, Pedicythere spp. 以及Henryhowella asperrima)的百分含量也相应发生了显著变化(图4,表3)。5个时期中,各属种的分布及其与水团变迁的关系简要描述如下。
表 3 B84A岩芯中介形虫主要属种指示的水团及其在各时代的百分含量Table 3. Correspondence of ostracode taxa with water masses, and their relative frequencies (%) in different times as recorded in core B84A种名 对应的水团 MIS 13–12 MIS 11–10 MIS 9–6 MIS 5 MIS 4–1 Polycope spp. AIW 44.9 7.7 66.4 34.1 42.8 Cytheropteron sedovi AODW下部 40.6 71.7 15.9 40.1 11.4 Microcythere medistriatum AIW&AODW 0 0 1.3 1.1 4.0 Pseudocythere caudata AIW 0 0 2.6 0.9 0.9 Pedicythere spp. AIW 0 0.5 0.5 1.7 0.9 Henryhowella asperrima AODW上部 0 0 0 0 24.0 Cytheropteron scoresbyi AODW&AIW 4.3 12.8 4.4 5.4 3.1 Cytheropteron higashikawai AODW下部 0 1.4 0.7 1.8 4.4 MIS 13–12期(约533~424 ka):Polycope spp.和Cytheropteron sedovi百分含量皆约为40%,相当于这两个属种在现代北冰洋2 000~3000 m水深(即AODW上部)的含量。这一时期,AODW & AIW指示种Cytheropteron scoresbyi也出现,但是含量较低(平均4.3%)。
MIS 11–10期(约424~337 ka):Cytheropteron sedovi丰度升至60%~90%,而Polycope spp.降至约20%,相当于这两个属种在现代北冰洋>3000 m水深(即AODW下部)的百分含量。这一时期Cytheropteron scoresbyi的含量也较高(平均12.8%)。
MIS 9–MIS 5早期(约337~100 ka):Cytheropteron sedovi降至10%~20%,Polycope spp. 升至60%~80%,相当于处在现代北冰洋1000~2 000 m的AIW中。另一个AIW指标种Pseudocythere caudata在这一时期的含量也达到最高(平均2.6%)。
MIS 5中–晚期(约100~71 ka):Polycope从>60%降到不足10%,而Cytheropteron sedovi从约10%增加到90%,显示AODW下部(>3000 m)的水团快速上涌。这一时期里,AIW指示种Pseudocythere caudata(平均0.9%)和Pedicythere spp.(平均1.7%),AIW & AODW指示种Microcythere medistriatum(平均1.1%)和Cytheropteron scoresbyi(平均5.4%),以及AODW下部指示种Cytheropteron higashikawai(平均1.8%)也出现,但是数量都很少。
楚科奇海台介形虫化石群的记录显示,MIS 5中–晚期时,在西北冰洋中层水域(<1000 m)曾发生过水团上涌事件:下方的AIW向上迁移,占据了原本属于AW的空间[17]。本研究则揭示了同一时期北冰洋中部更深水域(>2 000 m)的水团变迁:AODW向上迁移,将盘踞于此的AIW排挤了出去。MIS 5中–晚期的水团上涌事件,在罗蒙诺索夫脊以西的中层和深层水域可能是联动发生、同时进行的。本研究结果丰富了我们对于北冰洋深层水团变迁的认知,同时也印证了此前的研究结论[17]的可信度。
MIS 4–MIS 1期(约71 ka至今):Polycope spp. 含量逐渐恢复到20%~60%,Cytheropteron sedovi则降到约20%,相当于处在现代北冰洋2 000~3000 m的 AODW上部。这一时期AODW上部指示种Henryhowella asperrima首次出现并且其含量达到了很高值(平均24.0%)。
综上,本研究通过分析来自阿尔法脊的一根岩芯中的介形虫化石群,重建了北冰洋中部深层水团自MIS 13期以来所经历的下沉—上涌—下沉的变迁过程,同时也揭示了古海冰消长和古生产力变迁历史。本研究再次证明介形虫是北冰洋古海洋环境的有用指标,研究结论有待更多的北冰洋微体化石记录以及地球化学指标来进一步证实。
4. 结论
(1)由介形虫所代表的底栖生物古生产力在MIS 13–10期很低,进入MIS 9期之后显著升高;在MIS 9期之后,古生产力随气候冷暖交替而增减,间冰期的生产力通常显著高于冰期。
(2)海冰指示种Acetabulastoma arcticum在B84A岩芯中的分布,表明常年海冰很可能是在MIS 9期之后才出现的。
(3)基于AIW指示种Polycope spp.和AODW指示种Cytheropteron sedovi百分含量而重建的阿尔法脊约2280 m水深处水团变迁历史如下:MIS 13–12期,处于AODW上部;MIS 11–10期,AODW下部上涌,取代了AODW上部;MIS 9–MIS 5早期,上方的AIW下潜,取代了AODW;MIS 5中–晚期,AODW下部快速上涌,将AIW排挤出去;最终在MIS 4–1期,水团定格在AODW上部。其他几个常见种的百分含量变化与上述结论互不矛盾或互为印证。
致谢:本研究使用的岩芯由“雪龙号”科考船所采取。中国极地研究中心样品库为本研究提供了岩芯样品。感谢同济大学张灵敏博士在SEM照片拍摄过程中给予的指导;感谢上海交通大学章陶亮博士在绘制本文插图时给予的帮助。
-
![]()
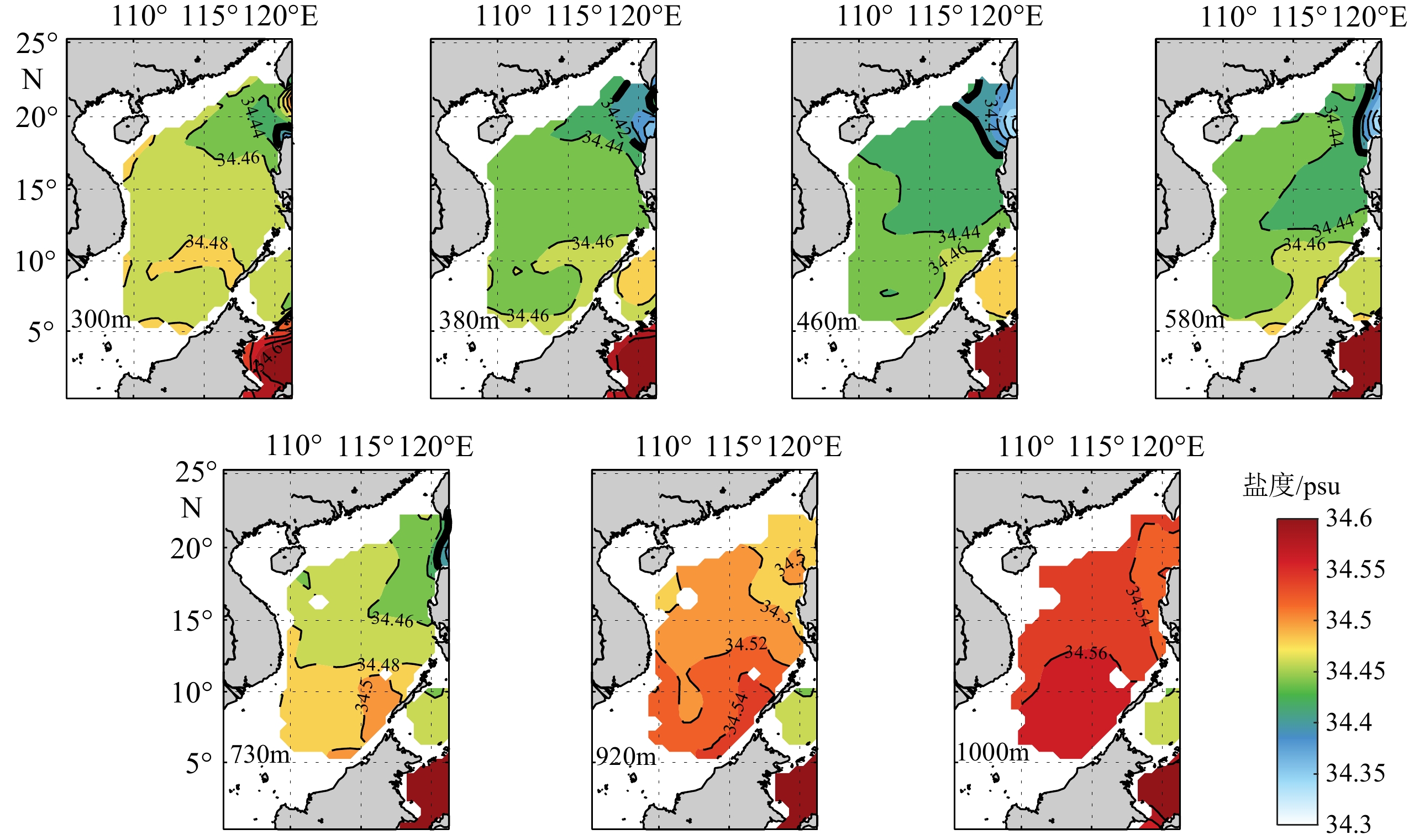
图 1 南海中层水的分布
使用SODA数据集,展示不同深度层中层水的分布范围,可见460 m层中层水的低盐特征最为突出。
Figure 1. The distribution of the intermediate water in the South China Sea
The SODA datasets present the distribution range of the intermediate water at different depths, and the low-salinity is the most prominent in the 460 m layer.
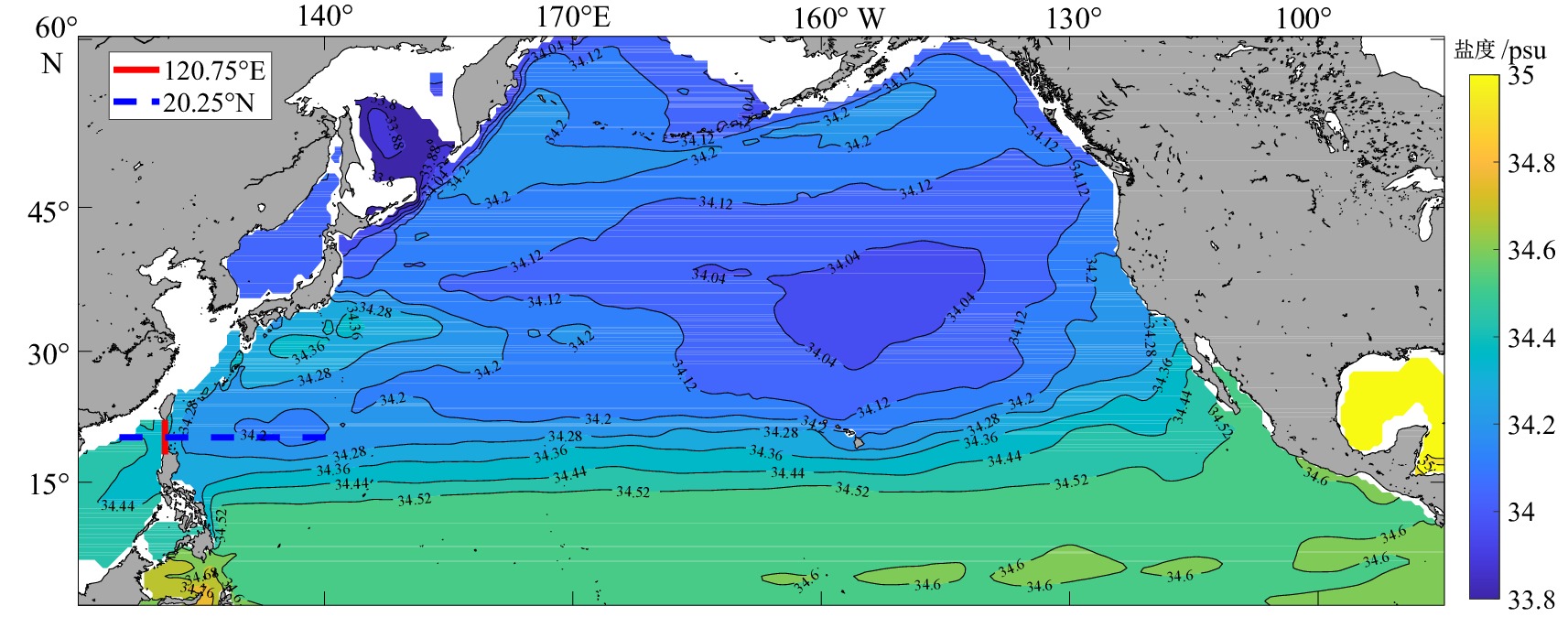
![]()
图 2 研究断面位置图
填色为580 m层盐度值。
Figure 2. Location map of the transection studied
Fill in the salinity value of the 580 m.
![]()
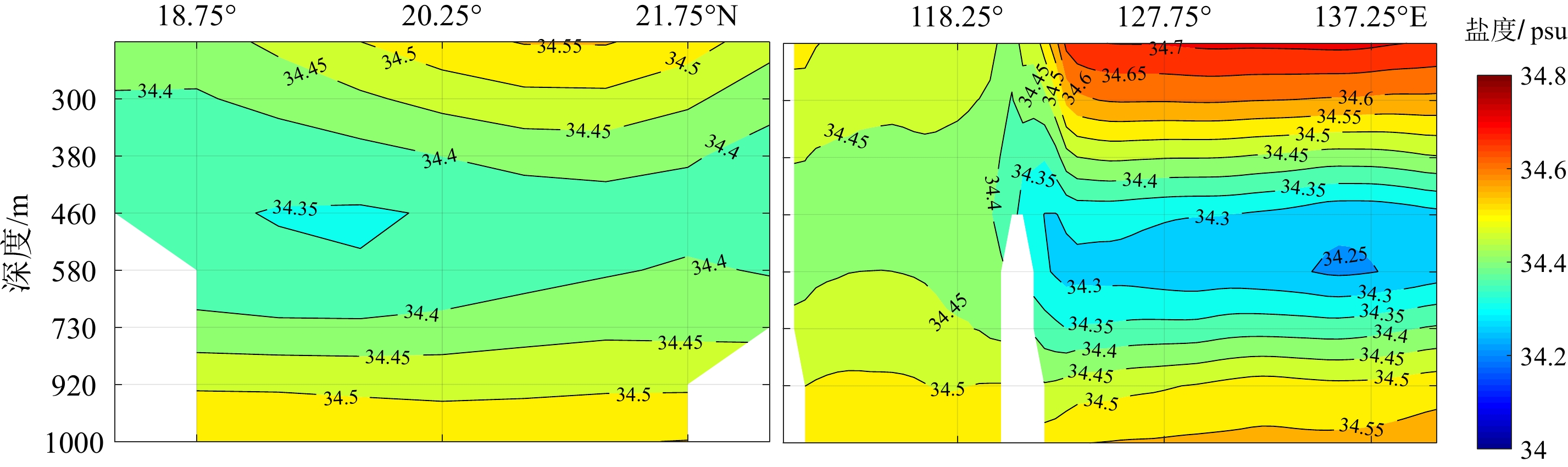
图 3 140 a南海纵断面盐度的平均状况分布图
左:120.75°E断面;右:20.25°N断面。
Figure 3. Distribution of salinity in the South China Sea in the last 140 years
left: 120.75°E transection; right: 20.25°N transection.
![]()
图 4 南海北部中层水核心层(460 m处)的盐度月平均分布特征
Figure 4. Characteristics of monthly distribution of salinity at 460 m of the intermediate water in the northern South China Sea
![]()
图 5 18.25°N断面南海中层水平均月际变化
Figure 5. The monthly variation of the intermediate water in the South China Sea at 18.25°N
![]()
图 6 中层水盐度的年际变化曲线
Figure 6. Interannual variation in salinity of the intermediate water in South China Sea
![]()
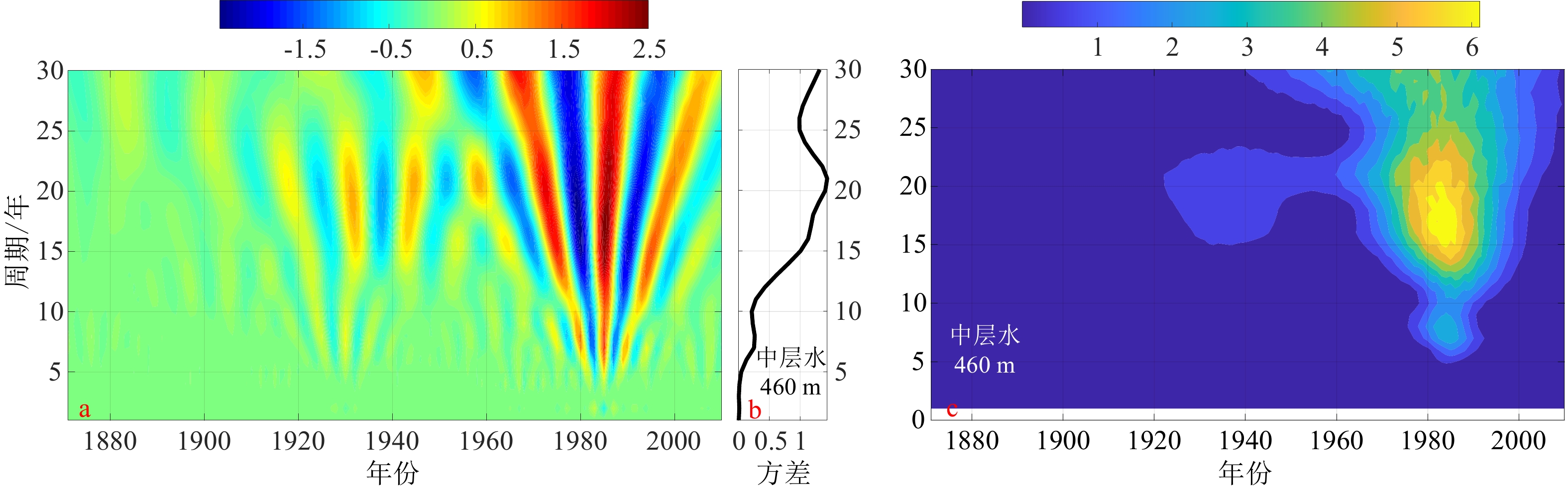
图 7 南海北部中层水盐度变化的年际周期小波分析图
a. 实部,b. 方差,c. 模方。
Figure 7. Wavelet analysis of the interannual cycle of salinity in the intermediate water of the South China Sea
a. The real part of wavelet coefficients, b. the variance, c. the modulus.
![]()
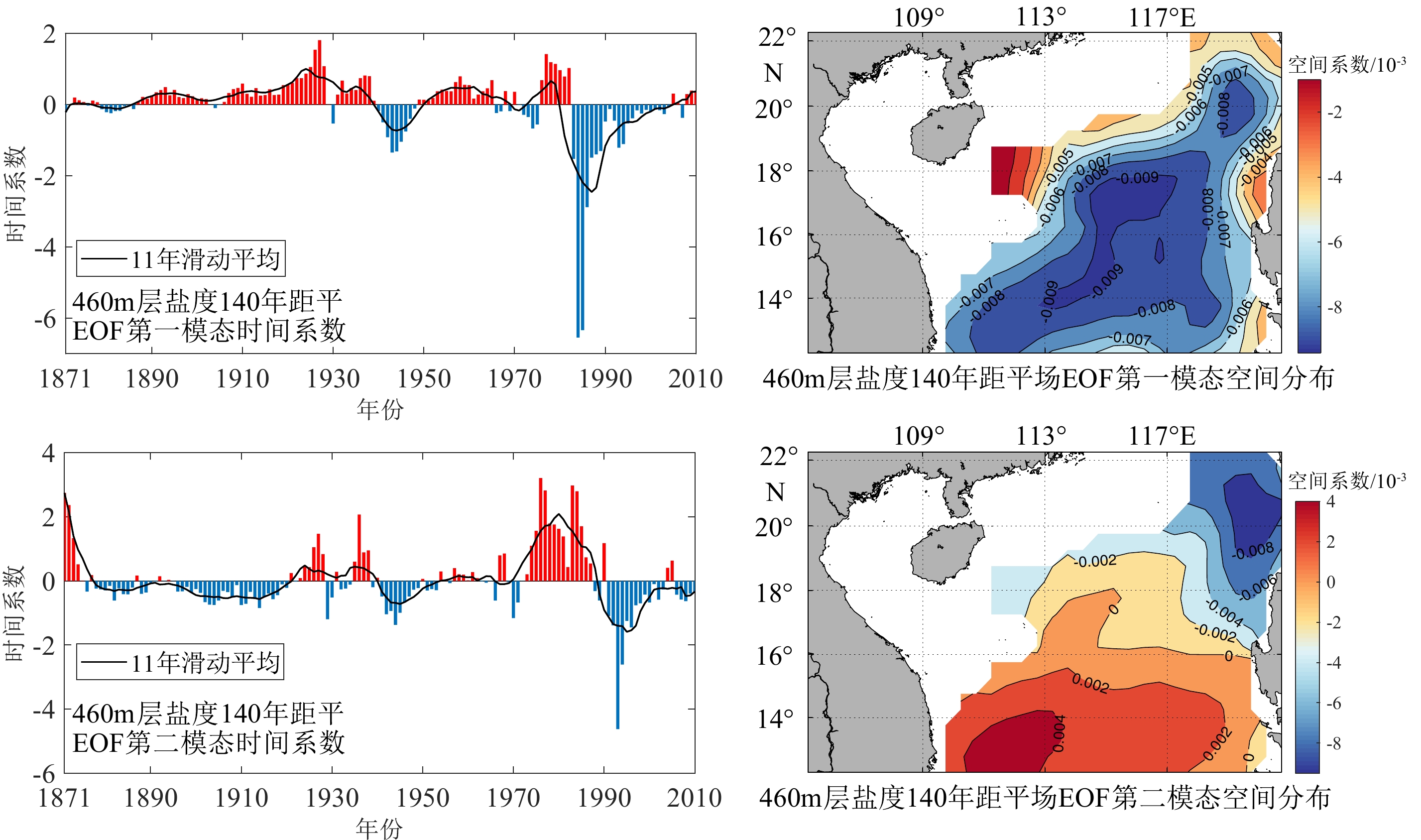
图 8 南海北部中层水盐度的年际模态分析
Figure 8. Interannual modal analysis on the salinity of the intermediate water in the northern South China Sea
![]()
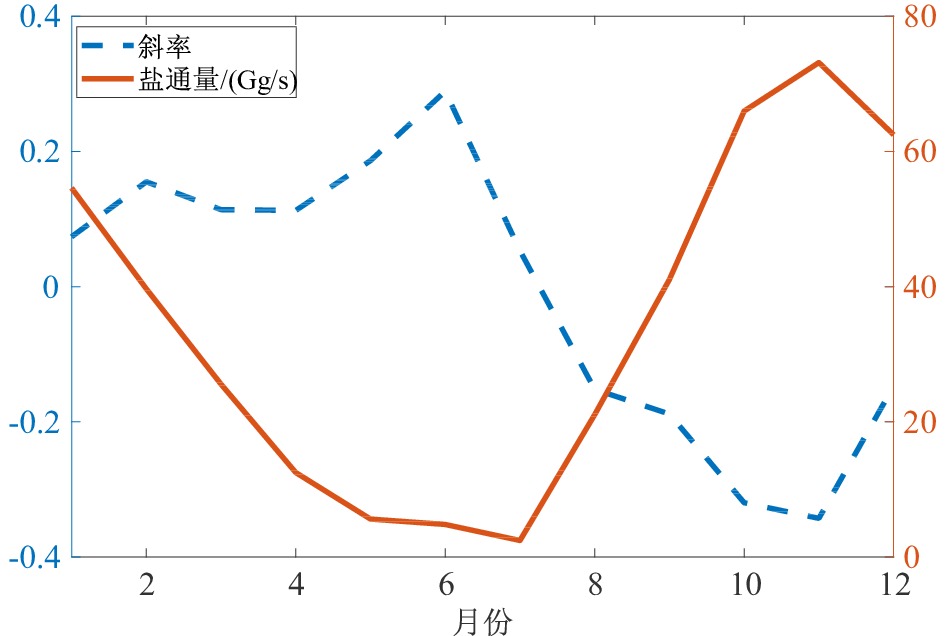
图 9 南海北部中层水120.75°E断面各月盐通量及变化率
Figure 9. Monthly salt flux and variation at 120.75°E transection of the intermediate water in the South China Sea
![]()
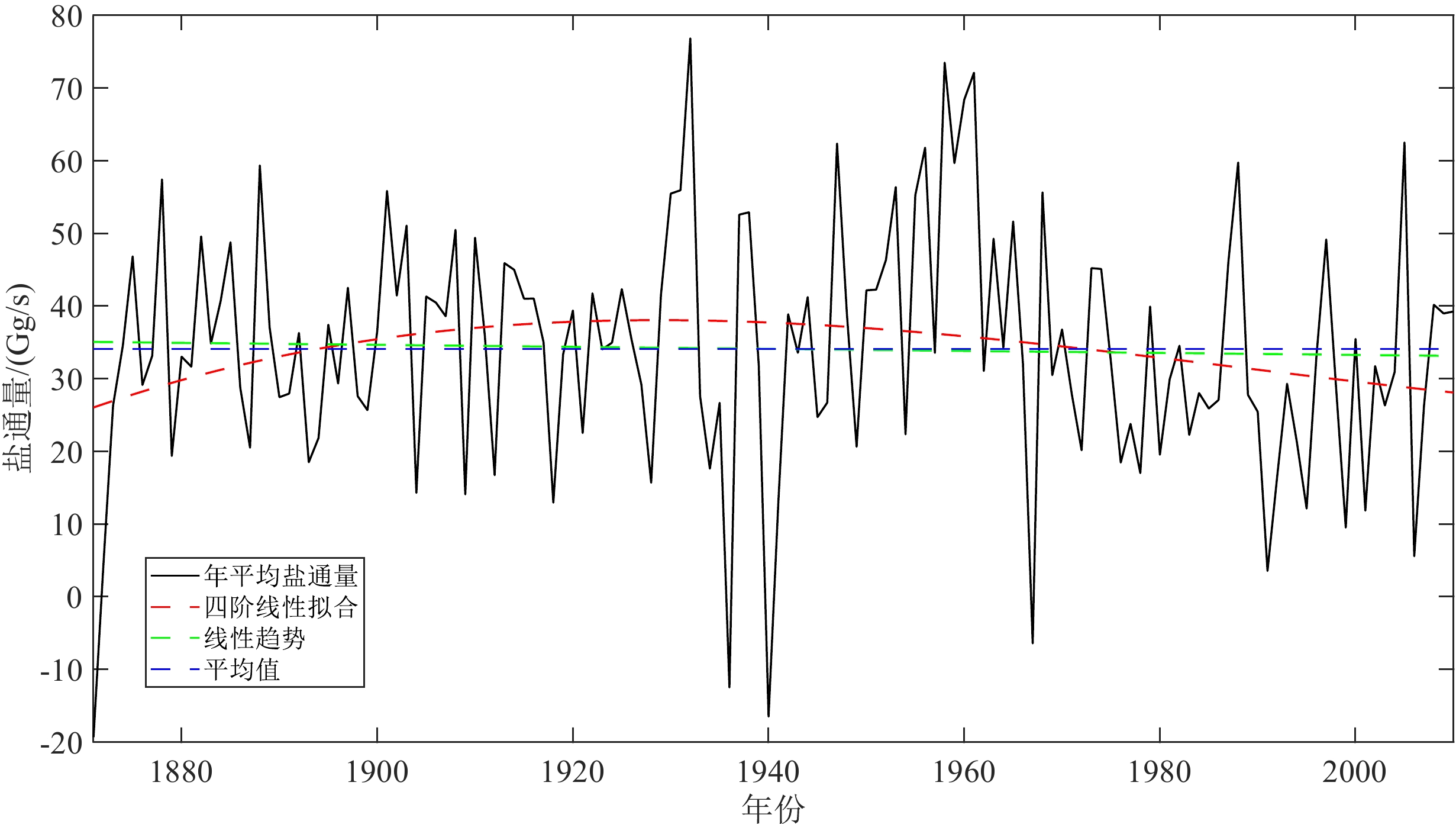
图 10 南海北部中层水盐通量的年际变化曲线
Figure 10. The interannual variation of water-salt flux in the intermediate water of the South China Sea
表 1 小波系数的实部正(负)位相年份与ENSO事件的对应
Table 1 The years of maximum (minimum) value of the real part of wavelet coefficients vs ENSO events
年份 小波实部正/负相位 ENSO事件 起止年月 峰值时间 强度 1965年 负相位 冷事件 1965.05—1966.05 1965.11 中等 1972年 正相位 冷事件 1972.05—1973.03 1972.11 强 1979年 负相位 冷事件 1979.09—1980.01 1980.01 弱 1986年 正相位 冷事件 1986.08—1988.02 1987.08 中等 1992年 负相位 冷事件 1991.05—1992.06 1992.01 中等 1995年 正相位 冷事件、
暖事件1994.09—1995.03、1995.09—1996.03 1994.12、1995.11 中等、弱  下载: 导出CSV
下载: 导出CSV
表 2 南海中层海水各模态贡献率
Table 2 Contribution of each mode to the intermediate seawater salinity of the South China Sea
第一模态 第二模态 第三模态 第四模态 贡献率 0.5973 0.1735 0.0533 0.0397
下载: 导出CSV
-
[1] Qu T D, Mitsudera H, Yamagata T. Intrusion of the North Pacific waters into the South China Sea [J]. Journal of Geophysical Research:Oceans, 2000, 105(C3): 6415-6424. doi: 10.1029/1999JC900323
[2] You Y Z, Chern C S, Yang Y, et al. The South China Sea, a cul-de-sac of North Pacific intermediate water [J]. Journal of Oceanography, 2005, 61(3): 509-527. doi: 10.1007/s10872-005-0059-6
[3] 刘增宏, 许建平, 孙朝辉, 等. 吕宋海峡附近海域水团分布及季节变化特征[J]. 热带海洋学报, 2011, 30(1):11-19 doi: 10.3969/j.issn.1009-5470.2011.01.002 LIU Zenghong, XU Jianping, SUN Chaohui, et al. The characteristics of water mass distribution and its seasonal variation near the Luzon Strait [J]. Journal of Tropical Oceanography, 2011, 30(1): 11-19. doi: 10.3969/j.issn.1009-5470.2011.01.002
[4] 乔方利. 中国区域海洋学-物理海洋学[M]. 北京: 海洋出版社, 2012. QIAO Fangli. Regional Oceanography of China Seas[M]. Beijing: China Ocean Press, 2012.
[5] 范立群, 苏育嵩, 李凤岐. 南海北部海区水团分析[J]. 中国海洋大学学报, 1989(S1):169-180 FAN Liqun, SU Yusong, LI Fengqi. Analysis of water masses in the northern area of the South China Sea [J]. Periodical of Ocean University of China, 1989(S1): 169-180.
[6] 李凤岐, 苏育嵩. 海洋水团分析[M]. 青岛: 青岛海洋大学出版社, 2000. LI Fengqi, SU Yusong. Ocean Water Mass Analysis[M]. Qingdao: Qingdao Ocean University Press, 2000.
[7] 田天, 魏皓. 南海北部及巴士海峡附近的水团分析[J]. 中国海洋大学学报, 2005, 35(1):9-12 TIAN Tian, WEI Hao. Analysis of water masses in the northern South China Sea and Bashi Channel [J]. Periodical of Ocean University of China, 2005, 35(1): 9-12.
[8] 李凤岐, 谢骏, 俎婷婷. 大洋性次-中层混合水团隶属函数的拟合及南海水团分析[J]. 中国海洋大学学报, 2004, 34(5):861-866 LI Fengqi, XIE Jun, ZU Tingting. Fitting method of membership function of the oceanic subsurface-intermediate mixing water mass and analysis of water masses in the South China Sea [J]. Periodical of Ocean University of China, 2004, 34(5): 861-866.
[9] Liu C J, Wang D X, Chen J, et al. Freshening of the intermediate water of the South China Sea between the 1960s and the 1980s [J]. Chinese Journal of Oceanology and Limnology, 2012, 30(6): 1010-1015. doi: 10.1007/s00343-012-1280-y
[10] Qu T D. Evidence for water exchange between the South China Sea and the Pacific Ocean through the Luzon Strait [J]. Acta Oceanologica Sinica, 2002, 21(2): 175-185.
[11] 韩钦臣, 康建成, HAN Guoqi, 等. 基于海洋分析资料的吕宋海峡水交换的月际变化特征[J]. 地球科学进展, 2015, 30(5):609-619 doi: 10.11867/j.issn.1001-8166.2015.05.0609 HAN Qinchen, KANG Jiancheng, HAN Guoqi, et al. Monthly variation characteristics of water exchange based on the analyzed ocean data in the Luzon Strait [J]. Advances in Earth Science, 2015, 30(5): 609-619. doi: 10.11867/j.issn.1001-8166.2015.05.0609
[12] Chen C T A, Huang M H. A mid-depth front separating the South China Sea water and the Philippine Sea water [J]. Journal of Oceanography, 1996, 52(1): 17-25. doi: 10.1007/BF02236530
[13] Chen C T A, Wang S L. Influence of intermediate water in the western Okinawa Trough by the outflow from the South China Sea [J]. Journal of Geophysical Research:Oceans, 1998, 103(C6): 12683-12688. doi: 10.1029/98JC00366
[14] Tian J W, Yang Q X, Liang X F, et al. Observation of Luzon Strait transport [J]. Geophysical Research Letters, 2006, 33(19): L19607. doi: 10.1029/2006GL026272
[15] Yang Q X, Tian J W, Zhao W. Observation of Luzon Strait transport in summer 2007 [J]. Deep Sea Research Part I:Oceanographic Research Papers, 2010, 57(5): 670-676. doi: 10.1016/j.dsr.2010.02.004
[16] 刘长建, 杜岩, 张庆荣, 等. 南海次表层和中层水团年平均和季节变化特征[J]. 海洋与湖沼, 2008, 39(1):55-64 doi: 10.3321/j.issn:0029-814X.2008.01.009 LIU Changjian, DU Yan, ZHANG Qingrong, et al. Seasonal variation of subsurface and intermediate water masses in the South China Sea [J]. Oceanologia et Limnologia Sinica, 2008, 39(1): 55-64. doi: 10.3321/j.issn:0029-814X.2008.01.009
[17] 王东晓, 刘雄斌, 王文质, 等. 理想海底地形的南海海洋经向翻转数值模拟[J]. 科学通报, 2004, 49(7):740-746 doi: 10.3321/j.issn:0023-074X.2004.05.014 WANG Dongxiao, LIU Xiongbin, WANG Wenzhi, et al. Simulation of meridional overturning in the upper layer of the South China Sea with an idealized bottom topography [J]. Chinese Science Bulletin, 2004, 49(7): 740-746. doi: 10.3321/j.issn:0023-074X.2004.05.014
[18] 王世红, 赵一丁, 尹训强, 等. 全球海洋再分析产品的研究现状[J]. 地球科学进展, 2018, 33(8):794-807 doi: 10.11867/j.issn.1001-8166.2018.08.0794 WANG Shihong, ZHAO Yiding, YIN Xunqiang, et al. Current status of global ocean reanalysis datasets [J]. Advances in Earth Science, 2018, 33(8): 794-807. doi: 10.11867/j.issn.1001-8166.2018.08.0794
[19] Carton J A, Giese B S. A reanalysis of ocean climate using simple ocean data assimilation (SODA) [J]. Monthly Weather Review, 2008, 136(8): 2999-3017. doi: 10.1175/2007MWR1978.1
[20] 方国洪, 魏泽勋, 崔秉昊, 等. 中国近海域际水、热、盐输运: 全球变网格模式结果[J]. 中国科学(D辑), 2003, 46(2):149-161 doi: 10.1360/03yd9014 FANG Guohong, WEI Zexun, CHOI C B, et al. Intercontinental water, heat and salt transport in China’s offshore waters: results from a global variable grid model [J]. Science in China Series D:Earth Sciences, 2003, 46(2): 149-161. doi: 10.1360/03yd9014
[21] 葛黎丽, 程旭华, 齐义泉, 等. 南海18°N断面上的体积和热盐输运[J]. 热带海洋学报, 2012, 31(1):10-17 GE Lili, CHENG Xuhua, QI Yiquan, et al. Upper-layer geostrophic volume, heat and salt transports across 18°N in the South China Sea [J]. Journal of Tropical Oceanography, 2012, 31(1): 10-17.
[22] Qiu C, Zhu J R. Assessing the influence of Sea level rise on salt transport processes and estuarine circulation in the Changjiang River estuary [J]. Journal of Coastal Research, 2015, 31(3): 661-670.
[23] 赵德平, 王卫强, 覃慧玲, 等. 南海18°N断面中层水的年代际变化[J]. 海洋学报, 2014, 36(9):56-64 ZHAO Deping, WANG Weiqiang, QIN Huiling, et al. Decadal changes of the intermediate water at 18°N in the South China Sea [J]. Acta Oceanologica Sinica, 2014, 36(9): 56-64.
[24] 苏纪兰. 南海环流动力机制研究综述[J]. 海洋学报, 2005, 27(6):1-8 SU Jilan. Overview of the South China Sea circulation and its dynamics [J]. Acta Oceanologica Sinica, 2005, 27(6): 1-8.
[25] . Qiao F L, Lü X G. Coastal upwelling in the South China Sea[M]//Satellite Remote Sensing of South China Sea. Taiwan: Tingmao Publish Company, 2008: 135-158.
[26] 李立, 吴日升, 郭小钢. 南海的季节环流: TOPEX/POSEIDON卫星测高应用研究[J]. 海洋学报, 2000, 22(6):13-26 LI Li, WU Risheng, GUO Xiaogang. Seasonal circulation in the South China Sea a TOPEX/POSEIDON satellite altimetry study [J]. Acta Oceanologica Sinica, 2000, 22(6): 13-26.
[27] 戴念军, 谢安, 张勇. 南海夏季风活动的年际和年代际特征[J]. 气候与环境研究, 2000, 5(4):363-374 doi: 10.3878/j.issn.1006-9585.2000.04.04 DAI Nianjun, XIE An, ZHANG Yong. Interannual and interdecadal variations of summer monsoon activities over South China Sea [J]. Climatic and Environmental Research, 2000, 5(4): 363-374. doi: 10.3878/j.issn.1006-9585.2000.04.04
[28] 赵永平, 陈永利. 南海暖池的季节和年际变化及其与南海季风爆发的关系[J]. 热带气象学报, 2000, 16(3):202-211 doi: 10.3969/j.issn.1004-4965.2000.03.002 ZHAO Yongping, CHEN Yongli. The seasonal and inter-annual variability of the South China Sea warm pool and its relation to the South China Sea monsoon onset [J]. Journal of Tropical Meteorology, 2000, 16(3): 202-211. doi: 10.3969/j.issn.1004-4965.2000.03.002
[29] 王雪竹. 南海中层水、中层环流及其与西北太平洋中层水交换研究[D]. 中国海洋大学硕士学位论文, 2010. WANG Xuezhu. The study of intermediate water, circulation in South China Sea and its intermediate water exchange with northwest pacific[D]. Master Dissertation of Ocean University of China, 2010.
[30] 冯晓伟, 孙照渤. ENSO对东亚夏季风强度的影响[J]. 南京气象学院学报, 1998, 21(2):196-200 FENG Xiaowei, SUN Zhaobo. The influnence of ENSO on the East Asian summer monsoon intensity [J]. Journal of Nanjing Institute of Meteorology, 1998, 21(2): 196-200.
[31] 朱益民, 杨修群, 陈晓颖, 等. ENSO与中国夏季年际气候异常关系的年代际变化[J]. 热带气象学报, 2007, 23(2):105-116 doi: 10.3969/j.issn.1004-4965.2007.02.001 ZHU Yimin, YANG Xiuqun, CHEN Xiaoying, et al. Interdecadal variation of the relationship between ENSO and summer interannual climate variability in China [J]. Journal of Tropical Meteorology, 2007, 23(2): 105-116. doi: 10.3969/j.issn.1004-4965.2007.02.001
[32] 徐霈强, 冯娟, 陈文. ENSO冷暖位相影响东亚冬季风与东亚夏季风联系的非对称性[J]. 大气科学, 2016, 40(4):831-840 doi: 10.3878/j.issn.1006-9895.1509.15192 XU Peiqiang, FENG Juan, CHEN Wen. Asymmetric role of ENSO in the link between the East Asian winter monsoon and the following summer monsoon [J]. Chinese Journal of Atmospheric Sciences, 2016, 40(4): 831-840. doi: 10.3878/j.issn.1006-9895.1509.15192






计量
- 文章访问数: 882
- HTML全文浏览量: 313
- PDF下载量: 107
