
Palynological evidence for palaeoenviromental change and human activity in Ningde of Fujian Province during Holocene
-
摘要: 分析了福建省宁德市东部海岸带地区NDQK2钻孔的42个孢粉样品。3个光释光和9个14C测年结果显示,钻孔沉积物年代跨越了末次冰消期晚期和全新世阶段。本文试图通过孢粉分析来重建宁德地区全新世以来的古环境演化,探讨古人类活动对天然植被演替的可能影响。本钻孔与该区域已发表的NDGK2和NDQK5孔的孢粉组合共同显示,深海氧同位素(MIS)5a阶段(约85 kaBP)至早全新世,周边陆地一直被亚热带常绿阔叶林和松林所覆盖,然而,约7770~4970 aBP期间以禾本科(Poaceae)为主的草本植物花粉和以芒萁属(Dicranopteris)为主的蕨类孢子含量剧增,孢粉组合与MIS5a以来的其他时段明显不同,类似于现代山地和海域表土孢粉组合。考虑到当地亚热带阔叶林被破坏后可形成以芒萁和禾本科为主的植被群落,我们认为宁德地区在7770~4970 aBP前就存在人类活动,并开始强烈影响森林组成,古人类砍伐树木导致分布于林缘与荒地的芒萁和禾本科等次生植被迅速扩张。基于东南沿海考古证据的时空分布对比,认为中、晚全新世本研究区周边古人类活动的出现可能与长江中下游古文明的迁移存在某种关联。Abstract: Analyzed by this paper are 42 pollen samples from the core of NDQK2, a 17.3 m long drilling core on the eastern coast of Ningde city, Fujian Province. Three optical stimulated luminescence (OSL) and nine AMS14C samples are dated. It is revealed that the core is made of with the deposits from Late Last Deglaciation and Holocene. Pollen are used as a major mean to reconstruct the Holocene palaeo-environment and to explore the possible impact of human activities on vegetation succession. Together with the published pollen data from adjacent cores of NDGK2 and NDQK5, it is found that subtropical evergreen broad-leaved forest and pine forest prevailed since marine isotope stage (MIS) 5a (~85 kaBP). However, Poaceae-dominated non-arboreal pollen component and Dicranopteris-dominated spores component increased sharply since 0.777~0.497 kaBP. The pollen assemblages are significantly different from other intervals of MIS5a, but similar to those in modern soils and marine sediments. If the expansion of Dicranopteris dichotoma and Poaceae plants is due to the destruction of subtropical broad-leaved forests by human activities, it suggests that human being had colonized in this region around 0.777~0.497 kaBP. The deforest by human being changed the natural forest composition, and resulted in the expansion of secondary plants such as Dicranopteris dichotoma and Poaceae distributed along the boarder of forest. The spatial and temporal distribution of archaeological evidence along southeast China indicates a possible link of human activities between the the study area and the migration of ancient culture from the lower Yangtze River during Mid-Late Holocene.
-
Keywords:
- pollen /
- human activities /
- the Holocene /
- Ningde City of Fujian Province
-
-
![]()
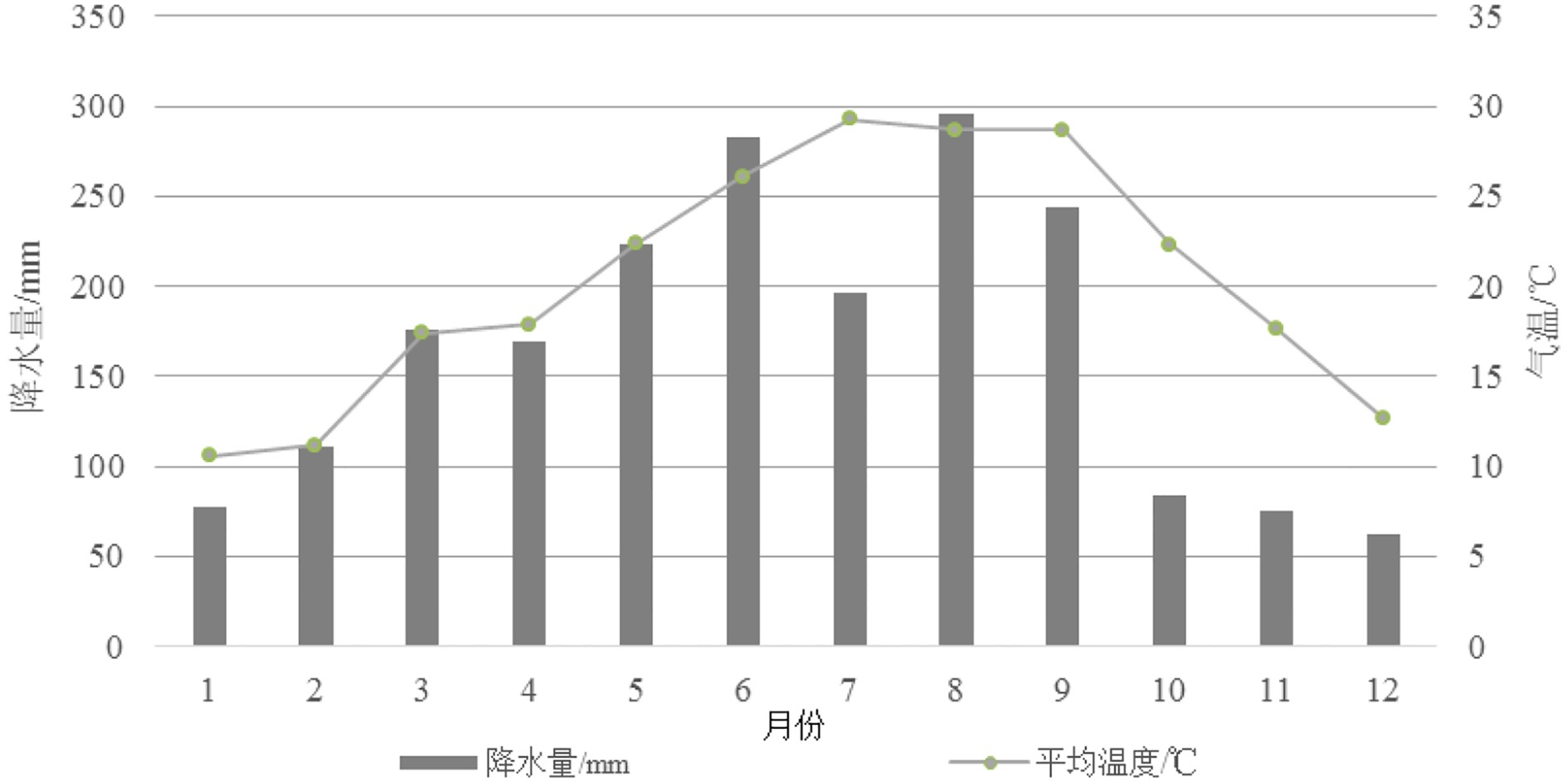
图 2 宁德月降水量与月均温趋势图(1981—2010年)
气候数据来源于中国气象数据网(http://data.cma.cn)。
Figure 2. Monthly precipitation and temperature in Ningde(1981—2010)
Climatic data are from China Meteorological Data Service Center(CMDC)(http://data.cma.cn).
![]()
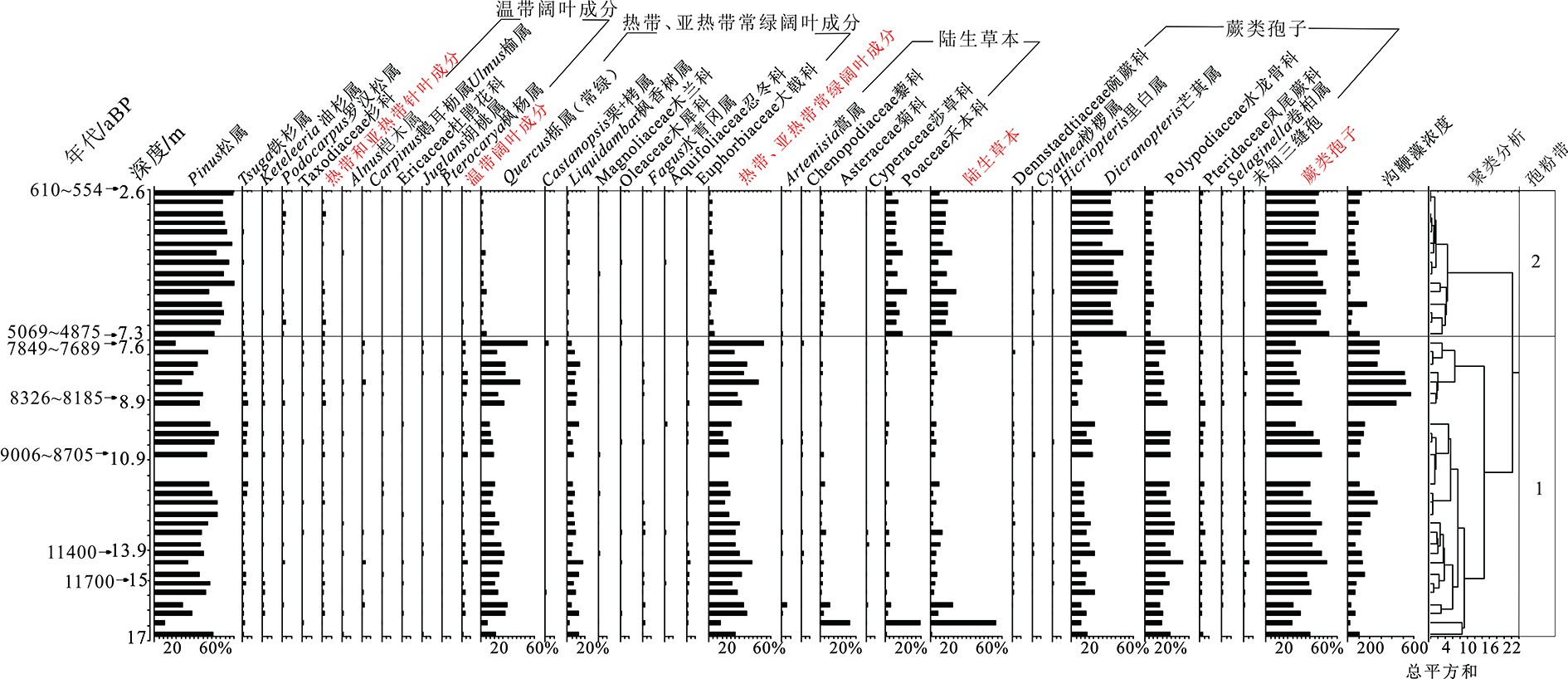
图 4 NDQK2钻孔主要花粉类型百分比及沟鞭藻浓度图
花粉百分比基于总花粉数量,孢子百分比基于花粉和孢子总数。
Figure 4. Percentage of major pollen taxa and concentration of marine dinocysts in core NDQK2
The percentage of pollen is based on the total number of pollen, and the percentage of spores is based on the total number of pollen and spores. The unit of the bottom number is %.
![]()
图 5 宁德表土花粉百分比图谱
花粉百分比基于总花粉数量,孢子百分比基于花粉和孢子总数。
Figure 5. Pollen percentage of surface samples in Ningde
The percentage of pollen is based on the total number of pollen, and the percentage of spores is based on the total number of pollen and spores. The unit of the bottom number is %.
![]()
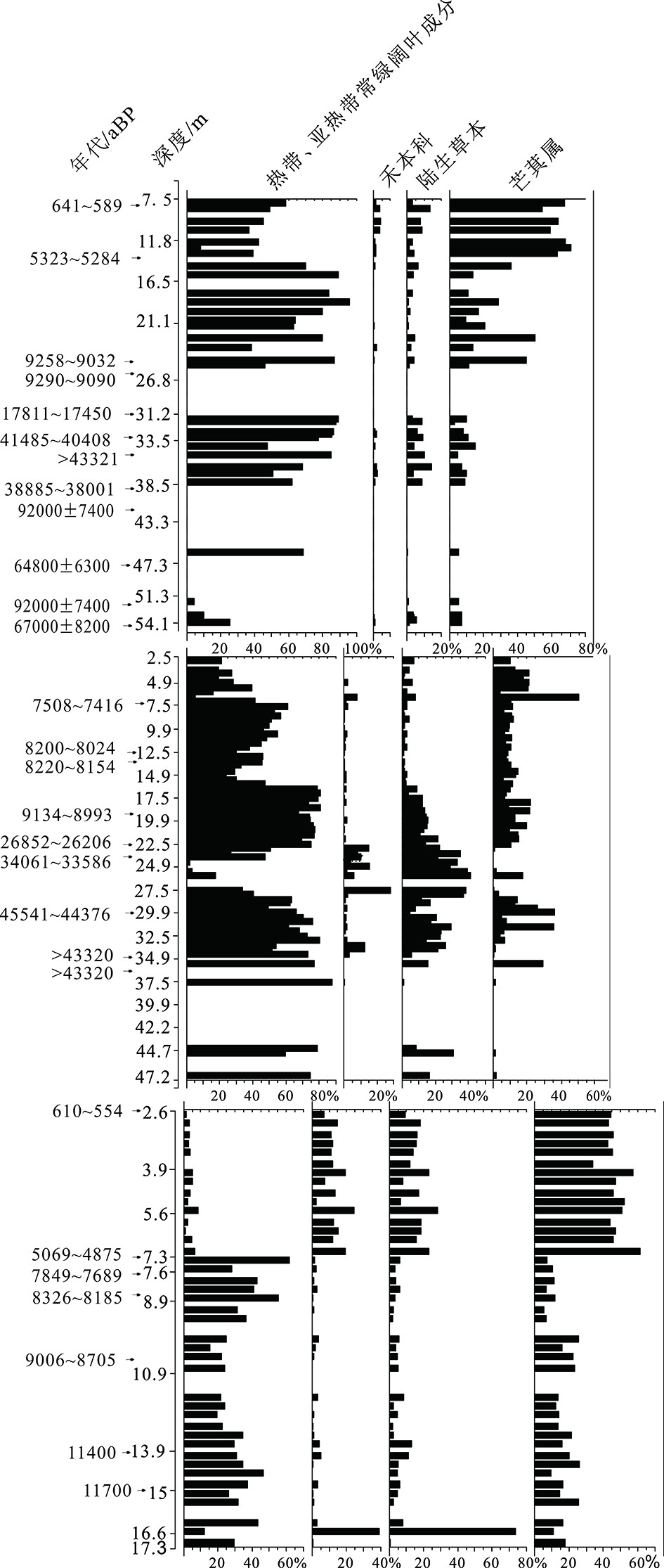
图 6 NDGK2、NDQK5和NDQK2孔中主要孢粉组的百分比对比图
花粉百分比基于总花粉数量,孢子百分比基于花粉和孢子总数。底部数字单位为%。NDGK2和NDQK5钻孔的孢粉数据来自文献[37]。
Figure 6. Comparison of percentages of pollen group in cores NDGK2, NDQK5 and NDQK2
The percentage of pollen is based on the total number of pollen, and the percentage of spores is based on the total number of pollen and spores. The unit of the bottom number is %. Pollen data from cores NDGK2 and NDQK5 are derived from ref.[37].
![]()
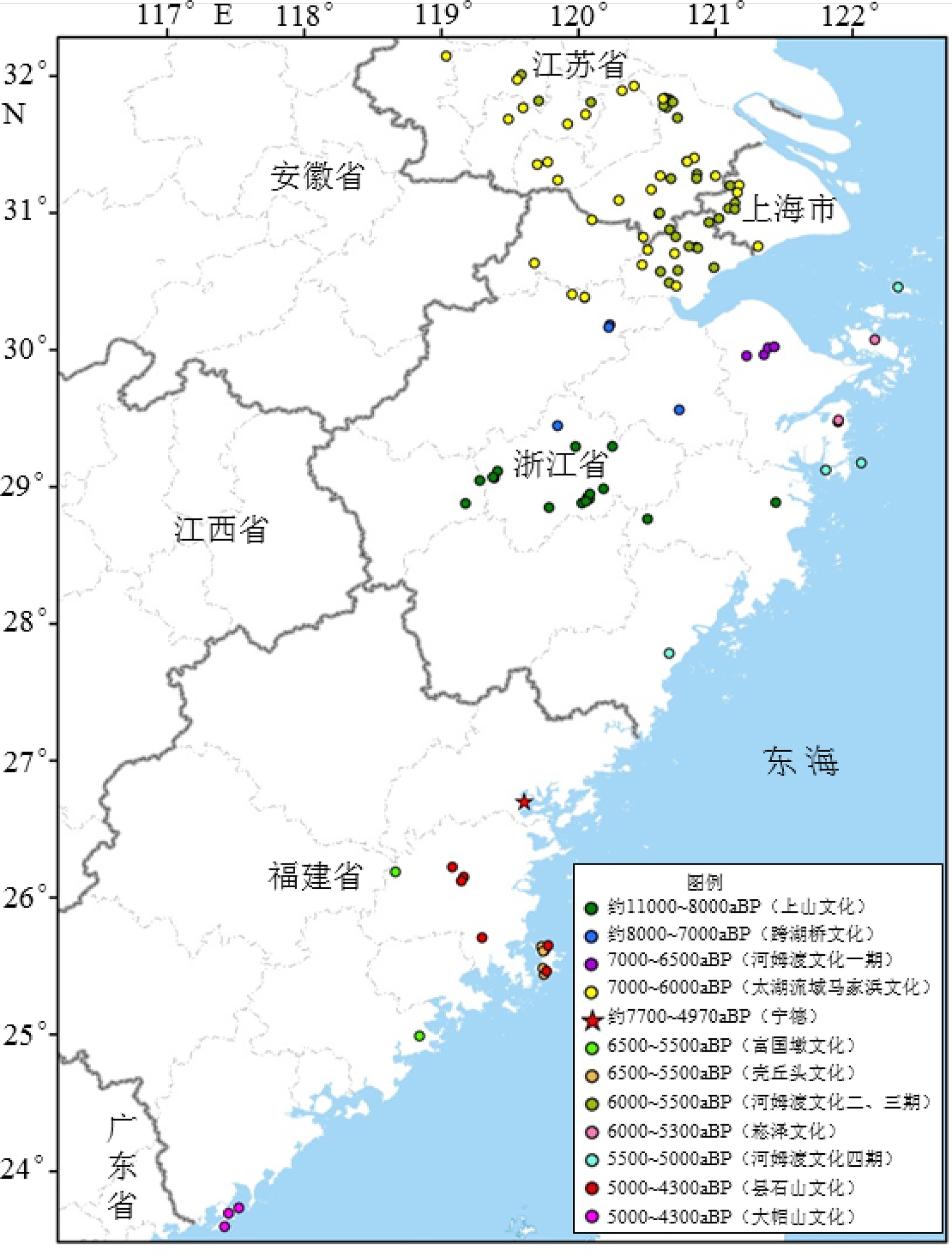
图 7 东南沿海11000~4300 aBP遗址时空分布图
Figure 7. Temporal and spatial distributions of archaeological sites between 11000 and 4300 aBP in the southeast coast of China
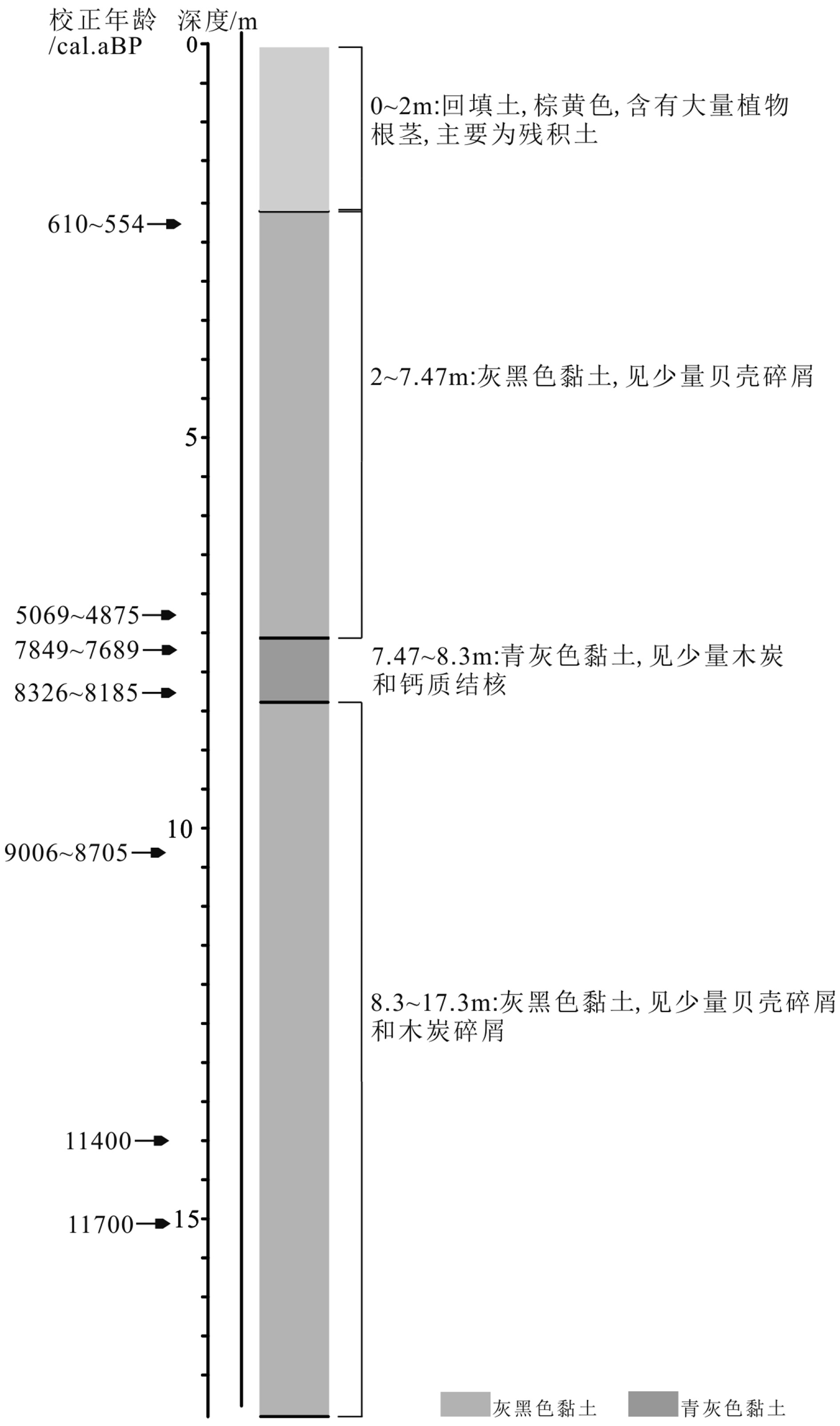
表 1 钻孔和现代表土采样点点位及样品类型描述
Table 1 Description of modern topsoil sampling points and sample types
样品编号 海拔/m 纬度 (N) 经度 (E) 样品类型 NDQK2钻孔 −2.6 26°38′09.79″ 119°33′04.28″ 黏土 1号 823.5 26°38′24.61″ 119°30′14.12″ 苔藓 12号 572 26°38′39.60″ 119°30′04.15″ 苔藓 15号 456 26°38′32.02″ 119°30′45.29″ 苔藓 64号 −7.3 26°41′54.02″ 119°39′52.01″ 泥 65号 −8.6 26°42′00.79″ 119°42′21.89″ 泥质砂 55号 −52.6 26°37′34.89″ 119°44′24.37″ 泥质砂  下载: 导出CSV
下载: 导出CSV
表 2 NDQK2钻孔的OSL测年结果
Table 2 OSL dated ages of core NDQK2
实验室编号 深度/m Th/ (μg/g) 误差/10−6 U (/μg/g) 误差/10−6 年龄/ka 误差/ ka 粒径/μm NDQK2-OSL-1 13.90~14.00 0.15 0.15 0.15 0.15 11.4 0.9 38~63 NDQK2-OSL-2 15.00~15.10 3.85 2.61 3.16 3.54 11.7 0.9 38~63 NDQK2-OSL-3 17.35~17.45 0.15 0.15 0.15 0.15 38~63
下载: 导出CSV
表 3 NDQK2钻孔的AMS14C测年结果
Table 3 AMS14C dating ages of core NDQK2
实验室编号 深度/m 测年材料 测试年龄/aBP 校正年龄 (2σ) /aBP 平均值/aBP NDQK2 14C-1 2.2 螺 625±20 610~554 582 NDQK2 14C-2 5.9 贝壳 465±20 531~500 515.5 NDQK2 14C-3 7.1 贝壳 505±20 543~510 526.5 Beta-567975 7.3 植物残体 4430±30 5069~4875 4972 Beta-552207 7.6 植物残体 6950±30 7849~7689 7769 NDQK2 14C-4 8.2 贝壳 7425±25 8326~8185 8255.5 NDQK2 14C-4-2 8.2 木炭 7055±30 7955~7835 7895 Beta-552202 10.3 植物残体 7990±40 9006~8705 8855.5 Beta-552203 13.3 植物残体 7970±30 8993~8704 8848.5
下载: 导出CSV
类型 主要成分 主要属种 孢粉 热带、亚热带针叶林成分 Podocarpus罗汉松属、Cupressaceae柏科、Keteleeria油杉属、Taxodiaceae杉科、Pinus松属 高海拔针叶成分 Abies冷杉属、Picea云杉属、Tsuga铁杉属 热带、亚热带阔叶成分 Quercus-evergreen常绿栎属、Castanopsis/Castanea栲属/栗属、Phyllanthus叶下珠属、Moraceae桑科、Myrica杨梅属、Acanthaceae爵床科、Theaceae山茶科、Rubiaceae茜草科、Sapindaceae无患子科、Hamamelidaceae金缕梅科、Liquidambar枫香树属、Verbenaceae马鞭草科、Ilex冬青属、Euphorbiaceae大戟科、Fagus水青冈属、Oleaceae木犀科、Rutaceae芸香科、Randia山黄皮属、Magnoliaceae木兰科 温带落叶成分 Quercus-decidious落叶栎属、Alnus桤木属、Ulmus榆属、Betula桦木属、Pterocarya枫杨属、Carpinus鹅耳枥属、Corylus榛属、Juglans胡桃属、Tilia椴属、Celtis朴属、Ericaceae杜鹃花科 草本植物 Artemisia蒿属、Chenopodiaceae藜科、Poaceae禾本科、Brassicaceae十字花科、Cyperaceae莎草科、Asteraceae菊科、Thalictrum唐松草属、Polygonaceae蓼科、Myriophyllum狐尾藻属、Typha香蒲属 蕨类 蕨类孢子 Polypodiaceae水龙骨科、Dicranopteris芒萁属、Pteris凤尾蕨属、Selaginella卷柏属、Hicriopteris里白属 藻类 淡水藻类 Concentricystes环纹藻、Pediastrum 盘星藻属 沟鞭藻囊孢 Operculodinium、Selenopemphix、Spiniferites、Tuberculodinium
下载: 导出CSV
表 5 我国东南沿海各期文化遗址信息
Table 5 Information of cultural relics in various stages along the southeast coast of China
下载: 导出CSV
-
[1] Flenley J R, King S M. Late Quaternary pollen records from Easter Island [J]. Nature, 1984, 307(5946): 47-50. doi: 10.1038/307047a0
[2] Diamond J M. The present, past and future of human-caused extinctions [J]. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 1989, 325(1228): 469-477. doi: 10.1098/rstb.1989.0100
[3] Cheng ZJ, Weng CY, Steinke S, et al. Anthropogenic modification of vegetated landscapes in southern China from 6,000 years ago [J]. Nature Geoscience, 2018, 11(12): 939-943. doi: 10.1038/s41561-018-0250-1
[4] 刘东生. 开展“人类世”环境研究, 做新时代地学的开拓者: 纪念黄汲清先生的地学创新精神[J]. 第四纪研究, 2004, 24(4):369-378 LIU Dongsheng. Demand of anthropocene study in the new stage of geoscience: in honor of late geologist Huang Jiqing for his innovative spirit [J]. Quaternary Sciences, 2004, 24(4): 369-378.
[5] Peng W X, Sonne C, Lam S S, et al. The ongoing cut-down of the Amazon rainforest threatens the climate and requires global tree planting projects: A short review [J]. Environmental Research, 2020, 181: 108887. doi: 10.1016/j.envres.2019.108887
[6] 白夜, 武达, 石宽, 等. 澳洲山火肆虐引发的思考[J]. 中国应急管理, 2020(1):62-63 BAI Ye, WU Da, SHI Kuan, et al. Reflections on the Raging bushfires in Australia [J]. China Emergency Management, 2020(1): 62-63.
[7] 王开发, 王宪曾. 孢粉学概论[M]. 北京: 北京大学出版社, 1982: 62-67. WANG Kaifa, WANG Xianzeng. Introduction to Palynology[M]. Beijing: Peking University Press, 1982: 62-67.
[8] 焦天龙. 东南沿海的史前文化与南岛语族的扩散[J]. 中原文物, 2002(2):13-16 JIAO Tianlong. Prehistoric culture in southeastern china and diffusion of Austronesian subgroups [J]. Cultural Relics of Central China, 2002(2): 13-16.
[9] 徐怡婷, 林舟, 蒋乐平. 上山文化遗址分布与地理环境的关系[J]. 南方文物, 2016(3):131-138 XU Yiting, LIN Zhou, JIANG Leping. The relationship between the distribution of Shangshan Culture Site and the geographical environment [J]. Cultural Relics in Southern China, 2016(3): 131-138.
[10] 王心喜. 长江下游原始文明新源头: 浙江嵊州小黄山新石器时代早期遗存的考古学研讨[J]. 文博, 2006(4):72-77 WANG Xinxi. A New source of primitive civilization in the lower reaches of the Yangtze River: Archaeological research on the early Neolithic relics in Xiao Huangshan in Shengzhou, Zhejiang [J]. Relics and Museology, 2006(4): 72-77.
[11] Zong Y, Chen Z, Innes J B, et al. Fire and flood management of coastal swamp enabled first rice paddy cultivation in east China [J]. Nature, 2007, 449(7161): 459-462. doi: 10.1038/nature06135
[12] 王海明, 刘淑华. 河姆渡文化的扩散与传播[J]. 南方文物, 2005(3):114-118, 113 WANG Haiming, LIU Shuhua. The spreading and propagation for Hemudu culture [J]. Cultural Relics in Southern China, 2005(3): 114-118, 113.
[13] 王斌. 马家浜文化研究[D]. 上海大学博士学位论文, 2019. WANG Bin. The comprehensive research of Majiabang culture[D]. Doctor Dissertation of Shanghai University, 2019.
[14] Yue Y F, Zheng Z, Rolett B V, et al. Holocene vegetation, environment and anthropogenic influence in the Fuzhou Basin, southeast China [J]. Journal of Asian Earth Sciences, 2015, 99: 85-94. doi: 10.1016/j.jseaes.2014.12.004
[15] 乐远福, 郑卓, ROLETT B V, et al. 闽江下游全新世孢粉记录的植被与环境变化[J]. 热带地理, 2016, 36(3):417-426 YUE Yuanfu, ZHENG Zhuo, ROLETT B V, et al. Vegetation and environmental changes inferred from a Holocene pollen spectrum in the Lower Reaches of Min River [J]. Tropical Geography, 2016, 36(3): 417-426.
[16] Yue Y F, Zheng Z, Huang K Y, et al. A continuous record of vegetation and climate change over the past 50, 000 years in the Fujian Province of eastern subtropical China [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 365-366: 115-123. doi: 10.1016/j.palaeo.2012.09.018
[17] Ma T, Zheng Z, Rolett B V, et al. New evidence for Neolithic rice cultivation and Holocene environmental change in the Fuzhou Basin, southeast China [J]. Vegetation History and Archaeobotany, 2016, 25(4): 375-386. doi: 10.1007/s00334-016-0556-0
[18] Zhao L, Ma C M, Leipe C, et al. Holocene vegetation dynamics in response to climate change and human activities derived from pollen and charcoal records from southeastern China [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 485: 644-660. doi: 10.1016/j.palaeo.2017.06.035
[19] 邱宏烈. 中国福建省亚热带山地4000年来植被变化的孢粉记录[J]. 亚热带资源与环境学报, 2006, 1(3):11-23 QIU Honglie. A 4000-Year pollen record of vegetation change from the subtropical mountains of Fujian Province, China [J]. Journal of Subtropical Resources and Environment, 2006, 1(3): 11-23.
[20] 吴少元. 中全新世以来福建东北地区植被与环境变化[J]. 微体古生物学报, 2019, 36(2):181-189 WU Shaoyuan. Reconstructing climate change and human actives based on pollen and spore data in northeast Fujian province since the middle Holocene [J]. Acta Micropalaeontologica Sinica, 2019, 36(2): 181-189.
[21] 林鹏, 丘喜昭, 吴志强, 等. 福建植被[M]. 福州: 福建科学技术出版社, 1990: 91-116. LIN Peng, QIU Xizhao, WU Zhiqiang, et al. Vegetation in Fujian[M]. Fuzhou: Fujian Science and Technology Press, 1990: 91-116.
[22] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980: 1-1375. WU Zhengyi. Chinese Vegetation[M]. Beijing: Science Press, 1980: 1-1375.
[23] Brown C A. Palynological Techniques[M]. 2nd ed. Dallas: AASP Foundation, 2008.
[24] 王伏雄, 钱南芬, 张玉龙, 等. 中国植物花粉形态[M]. 北京: 科学出版社, 1995. WANG Fuxiong, QIAN Nanfen, ZHANG Yulong, et al. Pollen Flora of China[M]. Beijing: Science Press, 1995.
[25] 唐领余, 毛礼米, 舒军武, 等. 中国第四纪孢粉图鉴[M]. 北京: 科学出版社, 2016: 1-620. TANG Lingyu, MAO Limi, SHU Junwu, et al. An Illustrated Handbook of Quaternary Pollen and Spores in China[M]. Beijing: Science Press, 2016: 1-620.
[26] Maher L J Jr. Statistics for microfossil concentration measurements employing samples spiked with marker grains [J]. Review of Palaeobotany and Palynology, 1981, 32(2-3): 153-191. doi: 10.1016/0034-6667(81)90002-6
[27] Reimer P J, Bard E, Bayliss A, et al. IntCal13 and Marine13 radiocarbon age calibration curves 0-50, 000 years cal BP [J]. Radiocarbon, 2013, 55(4): 1869-1887. doi: 10.2458/azu_js_rc.55.16947
[28] 陈金霞, 张德玉, 张文卿, 等. 末次冰期以来冲绳海槽北部古气候变化的孢粉记录[J]. 海洋学报, 2006, 28(1):85-91 CHEN Jinxia, ZHANG Deyu, ZHANG Wenqing, et al. The paleoclimatic change since the last glaciation in the north of Okinawa Trough based on the spore-pollen records [J]. Acta Oceanologica Sinica, 2006, 28(1): 85-91.
[29] Grimm E C. CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares [J]. Computers & Geoscience, 1987, 13(1): 13-35.
[30] 李宜垠, 周力平, 崔海亭. 人类活动的孢粉指示体[J]. 科学通报, 2008, 53(9):991-1002 doi: 10.1360/csb2008-53-9-991 LI Yiyin, ZHOU Liping, CUI Haiting. Palynological indicators of human activity [J]. Chinese Science Bulletin, 2008, 53(9): 991-1002. doi: 10.1360/csb2008-53-9-991
[31] 郑卓. 近几千年华南沿海地区植被的人为干扰[J]. 生态科学, 1998, 17(2):29-36 ZHENG Zhuo. Study of human disturbance to the vegetation in recent thousands of years in the coastal areas of southeast China [J]. Ecologic Science, 1998, 17(2): 29-36.
[32] 郑卓, 邓韫, 张华, 等. 华南沿海热带-亚热带地区全新世环境变化与人类活动的关系[J]. 第四纪研究, 2004, 24(4):387-393 doi: 10.3321/j.issn:1001-7410.2004.04.003 ZHENG Zhuo, DENG Yun, ZHANG Hua, et al. Holocene environmental changes in the tropical and subtropical areas of the South China and the relation to human activities [J]. Quaternary Research, 2004, 24(4): 387-393. doi: 10.3321/j.issn:1001-7410.2004.04.003
[33] Ma T, Rolett B V, Zheng Z, et al. Holocene coastal evolution preceded the expansion of paddy field rice farming [J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(39): 24138-24143. doi: 10.1073/pnas.1919217117
[34] 陈炽新, 邓韫, 郑卓, 等. 香港—大亚湾近岸海域表层沉积物的孢粉分布规律[J]. 热带海洋学报, 2004, 23(2):75-81 CHEN Zhixin, DENG Yun, ZHENG Zhuo, et al. Distribution characteristics of pollen and spore in surface sediments of nearshore waters between Hongkong and Daya Bay [J]. Journal of Tropical Oceanography, 2004, 23(2): 75-81.
[35] Huang K Y, Zheng Z, Liao W B, et al. Reconstructing late Holocene vegetation and fire histories in monsoonal region of southeastern China [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 393: 102-110. doi: 10.1016/j.palaeo.2013.11.005
[36] 罗传秀, 陈木宏, 刘建国, 等. 广东沿海及海南岛东南部海底表层孢粉分布及其环境意义[J]. 热带海洋学报, 2012, 31(2):55-61 doi: 10.3969/j.issn.1009-5470.2012.02.008 LUO Chuanxiu, CHEN Muhong, LIU Jianguo, et al. Pollen distribution in marine surface sediments of Guangdong coast and southeast Hainan Island and its environmental significance [J]. Journal of Tropical Oceanography, 2012, 31(2): 55-61. doi: 10.3969/j.issn.1009-5470.2012.02.008
[37] 于俊杰, 彭博, 兰佑, 等. 孢粉证据揭示MIS 5a以来福建东北沿海地区人类活动、海平面及气候变化[J]. 地球科学, 2019:1-23 YU Junjie, PENG Bo, LAN You, et al. Palynological record revealed anthropogenic deforestation, sea level and climate changes since marine isotope stage 5a in the northeastern coast of Fujian Province [J]. Earth Sciences, 2019: 1-23.
[38] 国家文物局. 中国文物地图集: 浙江分册[M]. 北京: 文物出版社, 2009. State Administration of Cultural Heritage. Atlas of Chinese Cultural Relics: Zhejiang Province[M]. Beijing: Cultural Relics Publishing House, 2009.
[39] 国家文物局. 中国文物地图集: 福建分册[M]. 北京: 文物出版社, 2007. State Administration of Cultural Heritage. Atlas of Chinese Cultural Relics: Fujian Province[M]. Beijing: Cultural Relics Publishing House, 2007.
[40] 高蒙河. 长江下游考古时代的环境研究: 文明化进程中的生态系统与人地关系[D]. 复旦大学博士学位论文, 2003. GAO Menghe. An environmental study on the Lower Reaches of the Yangtse River based on Archaeological Sequence: the ecological system and the relationship between human and environment in the process of civilization[D]. Doctor Dissertation of Fudan University, 2003.
[41] 邵九华. 河姆渡原始部落村[M]. 北京: 大众文艺出版社, 2008. SHAO Jiuhua. Hemudu Primitive Tribal Village[M]. Beijing: Popular Literature and Art Press, 2008.
[42] 周宗尧, 余国春, 董学发, 等. 河姆渡早期文化发展间断原因新考[J]. 中国地质, 2012, 39(2):550-558 doi: 10.3969/j.issn.1000-3657.2012.02.026 ZHOU Zongyao, YU Guochun, DONG Xuefa, et al. New study of the reasons for the interruption of the earliest Hemudu Culture [J]. Geology in China, 2012, 39(2): 550-558. doi: 10.3969/j.issn.1000-3657.2012.02.026
[43] 连琳琳, 张丽萍, 张妙仙, 等. 浙江沿海15226~5550 cal. aBP的古环境与海平面变化及其对河姆渡文化的影响[J]. 海洋地质与第四纪地质, 2014, 34(4):11-19 LIAN Linlin, ZHANG Liping, ZHANG Miaoxian, et al. Sporo-pollen Records of 15226~5550 cal. aBP on Zhejiang coast and implications for sea-level change [J]. Marine Geology & Quaternary Geology, 2014, 34(4): 11-19.
[44] 蔡保全. 从贝丘遗址看福建沿海先民的居住环境与资源开发[J]. 厦门大学学报: 哲社版, 1998(3):106-111 CAI Baoquan. A study of the living environment and resource development of the ancestors in coastal Fujian province from the Ruins of Beichu [J]. Journal of Xiamen University: Philosophy and Social Sciences Edition, 1998(3): 106-111.


计量
- 文章访问数: 2240
- HTML全文浏览量: 449
- PDF下载量: 92


