
High-resolution oxygen isotope records of Tridacna gigas from Palau, Western Pacific and its climatic and environmental implications
-
摘要: 砗磲是海洋中最大的双壳类贝壳,其碳酸盐壳体通常具有年纹层和天纹层,是一种理想的高分辨率古气候研究载体。氧同位素是砗磲古气候研究中最常用的指标之一,但在将其应用于古气候重建之前,通常需要对其现代地球化学过程进行准确的校准。帕劳群岛位于西太平洋暖池西北边缘,其珊瑚礁盘具有丰富的砗磲壳体资源,为开展古气候研究提供了丰富的材料。在本次研究中,对采自帕劳群岛的现代活体库氏砗磲(Tridacna gigas)PL-1的内层壳体进行了高分辨率氧同位素分析,同时利用该砗磲较为清晰的天生长纹层对氧同位素的年代学框架进行了标定。结果表明,该砗磲壳体的氧同位素没有明显的变化趋势,说明砗磲个体的生命效应对氧同位素没有显著影响;砗磲壳体氧同位素没有清晰的年周期变化,常出现不规则的毛刺状峰值。结合现代器测资料分析发现,帕劳砗磲内层壳体的氧同位素记录了热带太平洋ENSO活动对该区域水文气候变化的影响。该研究结果表明,帕劳砗磲内层壳体天生长纹层和氧同位素,具有用于开展高分辨率古气候研究的潜力。Abstract: Tridacna gigas is the largest marine bivalve, and its hard and dense aragonite shells usually have annual and daily growth lines, which have been demonstrated to be an ideal material for high-resolution paleoclimate research. The oxygen isotope has been widely used in Tridacna paleoclimate studies. However, the oxygen isotope of Tridacna shells must be accurately calibrated by modern geochemical process before paleoclimate reconstructions. Palau is located in the northwestern edge of the Western Pacific Warm Pool. Long-lived Tridacna spp. is a common species in the coral reefs of Palau Islands, which may provide abundant materials for paleoclimate reconstructions. In this study, we present a high-resolution oxygen isotope profile from the inner shell of a modern living T. gigas specimen PL-1 from Palau. The high-resolution chronology of the oxygen isotope profile is determined by the clear daily growth layers in the inner shell. The result suggests that the δ18Oc profile of the T. gigas shell has no obvious trend, indicating that the vital effects have no significant influence on the oxygen isotope of shell. Combining with the instrumental data, we found that the ENSO activities in the tropical Pacific had impacts on the regional hydro-climate changes of Palau, and left some fingerprint in the oxygen isotope of Tridacna shell. This study indicates that the daily growth layer and the oxygen isotope in the inner shell of Tridacna from Palau have the potential for high-resolution paleoclimate research.
-
Keywords:
- Tridacna spp. /
- daily growth layers /
- oxygen isotope /
- ENSO
-
长达200余万年的第四纪时期,洞庭湖区构造运动总趋势表现为凹陷,但其升降活动的幅度和强弱出现了多次变化。在湖区内部,受新构造运动差异凹陷与凸起的影响,洞庭盆地演变形成太阳山凸起、澧县凹陷、临澧凹陷、安乡凹陷、赤山凸起、华容凸起、沅江凹陷及湘阴凹陷等多个地质构造单元(图1),其活动主要受北东和西北向断裂的控制,并以北东向构造块体为主。西、南、东三面分别为武陵凸起、雪峰凸起和幕阜山凸起,北为江汉盆地。
![]() 图 1 洞庭盆地构造格局A1-武陵凸起,A2-雪峰凸起,A3-幕阜山凸起,A4-澧县凹陷,A5-临澧凹陷,A6-太阳山凸起,A7-安乡凹陷,A8-赤山凸起,A9-沅江凹陷,A10-华容凸起,A11-湘阴凹陷,A12-江汉盆地。Figure 1. Quaternary tectonic framework of Dongting BasinA1-Wuling Uplift,A2-Xuefeng Uplift,A3-Mufushan Uplift,A4-Lixian Sag,A5-Linli Sag,A6-Taiyangshan Uplift,A7-Anxiang Sag,A8-Chishan Uplift,A9-Yuanjiang Sag,A10-Huarong Uplift,A11-Xiangyin Sag,A12-Jianghan Basin.
图 1 洞庭盆地构造格局A1-武陵凸起,A2-雪峰凸起,A3-幕阜山凸起,A4-澧县凹陷,A5-临澧凹陷,A6-太阳山凸起,A7-安乡凹陷,A8-赤山凸起,A9-沅江凹陷,A10-华容凸起,A11-湘阴凹陷,A12-江汉盆地。Figure 1. Quaternary tectonic framework of Dongting BasinA1-Wuling Uplift,A2-Xuefeng Uplift,A3-Mufushan Uplift,A4-Lixian Sag,A5-Linli Sag,A6-Taiyangshan Uplift,A7-Anxiang Sag,A8-Chishan Uplift,A9-Yuanjiang Sag,A10-Huarong Uplift,A11-Xiangyin Sag,A12-Jianghan Basin.洞庭盆地构造沉积演化复杂,导致学者们对洞庭盆地第四纪地质问题的认识尚存在一定分歧。对于洞庭盆地第四纪构造属性,存在“断陷盆地”[1]、“拗陷盆地”[2-3]、“先断陷后拗陷”[4-5]等多种观点。此外,对近代洞庭湖历史演变也有不同见解,有人认为构造沉降是控制近代洞庭湖演变的关键因素[5-6],也有人认为泥沙淤积是控制近代洞庭湖演变的主要原因[7]。另外,对洞庭盆地出露的网纹红土成因,也有水成[8]和风成[9]两种观点。同时,也有学者对洞庭盆地第四纪构造沉积演变研究现状进行了总结论述[10-11]。但这些研究大都是针对整个洞庭湖第四纪,对盆地内不同构造单元的研究较少,其中柏道远等[12-18]分别对洞庭盆地安乡凹陷、澧县凹陷、华容凸起、临澧凹陷等内部构造单元的构造沉积特征与环境演化进行了深入研究;戴传瑞等[19-20]对洞庭盆地与周边造山带耦合关系及沅江凹陷地层沉积相特征及演化做了分析探讨。这些湖内不同构造单元第四纪地质研究,极大深化了洞庭盆地第四纪构造沉积演变特性研究。
第四纪以来,洞庭湖赤山凸起构造沉积演变活跃,致使其第四纪地层沉积时代、成因及构造活动较为复杂,而赤山凸起作为洞庭湖内部重要的地质构造单元,研究其第四纪构造沉积演变特性对探索洞庭湖第四纪历史演变有着重要的地质意义。
本文从地质构造、地层沉积、古气候、文化遗址等多角度入手,对洞庭湖赤山凸起第四纪沉积时代、成因及构造活动规律等进行深入探讨,以期丰富赤山凸起第四纪构造沉积演变特性研究成果。
1. 地质构造
根据洞庭盆地广布的第四纪堆积物及前人研究成果[10-11],第四纪期间,洞庭盆地各次级凹陷的构造活动总体为沉降,而周缘凸起区及盆地内部的赤山凸起总体为抬升,这一构造活动差异使湖区内部凹陷和周缘凸起区(包括赤山凸起)的第四纪沉积形成时代、成因、厚度、岩土体颗粒组成等特性具显著差异。
赤山凸起为洞庭盆地南部的小型凸起断块,主要受其东、西两侧正断裂所控制,东侧为大通湖-草尾断裂,西侧为目平湖断裂,形成一南北向地垒构造。大通湖-草尾断裂走向北北东,倾向东部沅江凹陷内部,其东盘下降,形成了下陷深达150 m以上的沅江凹陷,西盘上升,出露地层为古近纪红层、早更新世、中更新世及全新世地层,断距在150 m以上;目平湖断裂走向也为北北东,倾向西部安乡凹陷内部,其西盘下降,形成了安乡凹陷,东盘上升。
赤山凸起东西两侧发育了两条走向基本一致的反向倾斜的正断层,赤山凸起为中部地垒,其第四纪以来总体处于凸起状态,东西两侧则处于沉陷状态。
2. 地层沉积特性
赤山凸起大体为南北向展布的长条状低矮丘陵区,北端窄,仅有2.2 km,南端稍宽,为8~15 km,长约18 km,高程50~115 m,是洞庭湖内最大的岛屿,位于洞庭盆地南部,为东、西洞庭湖天然分界(图1),中部偏西为岛内地势最高地,东、南、北侧较低。其中较高地区域(基岩面海拔约为75 m)地表出露地层为早更新世汨罗组,次高地(基岩面海拔约为55 m)为中更新世新开铺组,低地(基岩面海拔约为0 m)为中更新世白沙井组,地势最高的中西部出露古近纪红层。
根据赤山凸起上已有钻孔资料、研究成果[13]及笔者长期地质调查测绘和钻探成果,赤山凸起地层沉积划分见表1。总体发育特征为:在赤山凸起南、北端基岩面海拔 0 m左右发育有更新世白沙井组、新开铺组、汨罗组和全新世地层,缺失晚更新世及中更新世晚期地层;高程55 m左右发育更新世新开铺组、汨罗组和全新世地层,缺失晚更新世及中更新世中、晚期地层;中西部基岩面海拔75 m左右发育少量早更新世汨罗组和全新世地层,缺失整个晚更新世和中更新世地层。3个不同高程基岩面区域第四纪沉积之间呈相互切割关系,组成镶嵌阶地(图2)。
表 1 赤山凸起第四纪地层划分Table 1. Quaternary stratigraphy of Chishan Uplift时代 名称 地层代号 厚度/m 全新世 全新世冲积 Qhal 3~10 晚更新世 缺失 中更新世 白沙井组 Qp2b 15~30 新开铺组 Qp2x 20~30 早更新世 汨罗组 Qp1m 5~8 ![]() 图 2 赤山凸起东西向地质剖面图A-赤山凸起南部新湾镇ZK1东西向地质剖面图,B-赤山凸起中部鸡婆村ZK2东西向地质剖面图。1-含碎石砂质黏土,2-砂质黏土,3-含卵砂质黏土,4-黏土,5-砂卵砾石,6-粉砂质黏土,7-泥质粉砂岩,8-第四系地层分界线,9-第四系与基岩分界线,10-钻孔编号及位置。Q4S-全新世人工堆积,Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 2. EW-trending geological profile through Chishan UpliftA-EW-trending geological section of ZK1 through Xinwan town in the south of Chishan Uplift,B- EW-trending geological section of ZK2 through Jipo village in the middle of Chishan Uplift.1-sandy clay with gravel,2-sandy clay,3-sandy clay with pebbles,4-clay,5-sand gravel,6-silty clay,7-pelitic siltstone,8-Quaternary stratigraphic boundary,9-boundary between Quaternary system and bedrock,10- number and location of drilling hole.Q4S-artificial accumulation of Holocene,Q4edl-wind accumulation of Holocene,Qp2b-Baishajing Formation of Middle Pleistocene, Qp2x-Xinkaipu Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene;E-Paleogene.
图 2 赤山凸起东西向地质剖面图A-赤山凸起南部新湾镇ZK1东西向地质剖面图,B-赤山凸起中部鸡婆村ZK2东西向地质剖面图。1-含碎石砂质黏土,2-砂质黏土,3-含卵砂质黏土,4-黏土,5-砂卵砾石,6-粉砂质黏土,7-泥质粉砂岩,8-第四系地层分界线,9-第四系与基岩分界线,10-钻孔编号及位置。Q4S-全新世人工堆积,Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 2. EW-trending geological profile through Chishan UpliftA-EW-trending geological section of ZK1 through Xinwan town in the south of Chishan Uplift,B- EW-trending geological section of ZK2 through Jipo village in the middle of Chishan Uplift.1-sandy clay with gravel,2-sandy clay,3-sandy clay with pebbles,4-clay,5-sand gravel,6-silty clay,7-pelitic siltstone,8-Quaternary stratigraphic boundary,9-boundary between Quaternary system and bedrock,10- number and location of drilling hole.Q4S-artificial accumulation of Holocene,Q4edl-wind accumulation of Holocene,Qp2b-Baishajing Formation of Middle Pleistocene, Qp2x-Xinkaipu Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene;E-Paleogene.2.1 赤山凸起基岩面海拔0 m左右区域地层特性
为研究赤山凸起基岩面海拔0 m左右区域地层特性,笔者在赤山岛南部新湾镇镇区204省道东部空地上布置了一个钻孔,该钻孔揭露第四系厚度为52.31 m,分为全新世、中更新世中期白沙井组和早期新开铺组以及早更新世晚期汨罗组地层,缺失中更新世晚期及晚更新世地层,其第四纪地层自上而下可分为8层(图3):
![]() 图 3 赤山岛南部新湾镇ZK1综合柱状图及南北向地质剖面图A-南部新湾镇ZK1综合柱状图,B-南部新湾镇ZK1南北向地质剖面图,Q4S-全新世人工堆积,Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 3. Integrated column of ZK1 and NS-trending geological profile through Xinwan town, Southern Chishan UpliftA-Comprehensive histogram of ZK1 in Xinwan town in the south,B-NS-trending geological section of ZK1 in Xinwan town in the south;Q4S-artificial accumulation of Holocene,Q4edl-wind accumulation of Holocene,Qp2b-Baishajing formation of Middle Pleistocene,Qp2x-Xinkaipu Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.
图 3 赤山岛南部新湾镇ZK1综合柱状图及南北向地质剖面图A-南部新湾镇ZK1综合柱状图,B-南部新湾镇ZK1南北向地质剖面图,Q4S-全新世人工堆积,Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 3. Integrated column of ZK1 and NS-trending geological profile through Xinwan town, Southern Chishan UpliftA-Comprehensive histogram of ZK1 in Xinwan town in the south,B-NS-trending geological section of ZK1 in Xinwan town in the south;Q4S-artificial accumulation of Holocene,Q4edl-wind accumulation of Holocene,Qp2b-Baishajing formation of Middle Pleistocene,Qp2x-Xinkaipu Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.①层以黄褐、灰褐色含碎石砂质黏土为主,并夹有生活垃圾和建筑垃圾等,厚度为3.74 m;为全新统人工堆积。
②层为红褐色含细粒砂质黏土,呈网纹状,明显经过湿热化,网纹细长而散乱无规则,长度约1~10 cm不等,厚度为3.22 m,河湖、湖泊相沉积,为中更新统白沙井组湖积堆积。
③层为白色—浅灰黄色粉砂质黏土,中间夹一层厚度为0.2 m砂层,硬塑状,结构密实,手摸岩芯有轻微砂感,厚度为2.13 m,河湖、湖泊相沉积,为中更新统白沙井组湖积堆积。
④层为黄赤带紫红色黏土,质纯,黏性、韧性强,硬塑状,结构密实,刀切面光滑,厚度为8.26 m,河湖、湖泊相沉积,为中更新统白沙井组湖积堆积。
⑤层为灰紫色粉砂质黏土,夹三层厚度分别约为0.1、0.4、0.5 m粉砂层和少量卵砾石,硬塑状,结构密实,手摸岩芯有轻微砂感,厚度为10.14 m,河湖相沉积,为中更新统新开铺组冲湖积堆积。
⑥层为灰红色粉砂质黏土,硬塑状,结构密实,手摸岩芯有轻微砂感,厚度为5.09 m,河湖相沉积,为早更新统汨罗组冲湖积堆积。
⑦层为黄褐色砂砾层,砾石占50%~65%,砂约占30%~40%,砾石成分为砂岩、硅质岩、板岩等,磨圆度一般较差,粒径一般小于3 cm,厚度约为18.82 m,河流、河湖相,略具定向排列,优势产状近似NE向。为早更新统汨罗组冲湖积堆积。
⑧层为红褐色泥质粉砂岩,古近纪地层,未揭露层底。
2.2 赤山凸起基岩面海拔55 m左右区域地层特性
笔者在赤山岛中部南嘴镇鸡婆村东南部岗地上也布置了一个钻孔,研究基岩面海拔55 m左右区域地层特性,揭露第四纪厚度为25.88 m,分别为早更新世晚期汨罗组和中更新世早期新开铺组地层,缺失晚更新世、中更新世中期和晚期地层,其第四纪地层自上而下可分为5层(图4):
![]() 图 4 赤山岛中部鸡婆村ZK2综合柱状图及南北向地质剖面图A-中部鸡婆村ZK2综合柱状图,B-中部鸡婆村ZK南北向地质剖面图。Q4edl-全新世风成堆积,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 4. Integrated column of ZK2 and NS-trending geological profile through Xinwan town, Southern Chishan UpliftA-Comprehensive histogram of ZK2 in the central section,B- NS-trending geological section of ZK2 in the central section.Q4edl-wind accumulation of Holocene,Qp2b-Baishajing Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.
图 4 赤山岛中部鸡婆村ZK2综合柱状图及南北向地质剖面图A-中部鸡婆村ZK2综合柱状图,B-中部鸡婆村ZK南北向地质剖面图。Q4edl-全新世风成堆积,Qp2x-中更新世新开铺组,Qp1m-早更新世汨罗组,E-古近纪。Figure 4. Integrated column of ZK2 and NS-trending geological profile through Xinwan town, Southern Chishan UpliftA-Comprehensive histogram of ZK2 in the central section,B- NS-trending geological section of ZK2 in the central section.Q4edl-wind accumulation of Holocene,Qp2b-Baishajing Formation of Middle Pleistocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.①紫红、红褐色粉砂质黏土,表层厚度约2.1 m,呈网纹状,明显经过湿热化,土中存在交错层理的砂层透镜体,网纹细长而散乱无规则,长度约1~12 cm不等,厚度为7.24 m,河湖相沉积,为中更新统新开铺组湖积堆积。
②灰褐色含砾粉砂质黏土,硬塑状,结构密实,手摸岩芯有轻微砂感,厚度为3.72 m,河湖相沉积,为早更新统汨罗组冲湖积堆积。
③暗紫红色砾石层夹砂砾层,砾石成分以石英砂岩为主,脉石英、硅质岩次之,填隙物为含细砾黏土质粗—细砂,弱固结,局部夹有黏土层透镜体,厚度为3.18 m,河流、河湖相沉积,为早更新统汨罗组冲湖积堆积。
④灰红色砾石层,砾石含量90%左右,成分中以脉石英和硅质岩为主,约80%,其他为石英砂岩,磨圆多较好,呈次圆—圆状,且具有上细下粗特征,粒径上部多为1~5 cm,下部多为4~20 cm,砾石间填隙物多为粗—细砂,含量约8%~14%,且该层上部夹有厚15 cm左右的砂层,厚度为11.36 m,河流、河湖相沉积,为早更新统汨罗组冲湖积堆积。
⑤红褐色泥质粉砂岩,为古近纪红层,未揭露层底。通过钻孔附近人工开挖露头剖面还发现,②层在赤山凸起东北部砂层透镜体过渡为砂层,且砂层逐渐增厚,此特性揭露了该地层沉积过程中赤山凸起区域存在着自南西往北东持续的水流运动;③层砾石略具定向,优势产状约为230°∠18°;④层砾石呈定向沉积特征更为明显,优势产状大致与③层相近,部分呈倾斜状切蚀下伏层,表明④层沉积过程中,赤山凸起区域内水流方向可能长期保持由SWW向NEE,且顺水流方向有由薄缓慢变厚的趋势。
2.3 赤山凸起基岩面海拔75 m左右区域地层特性
经过长期地质调查测绘,在赤山凸起中西部、基岩面海拔75 m左右区域第四系主要为全新统和早更新世晚期汨罗组地层,且部分区域汨罗组地层剥蚀较为严重,甚至已不完整或缺失,通过零星露头剖面,其第四纪沉积自上而下可分为5层(图5)。
![]() 图 5 赤山凸起基岩面海拔75 m左右南北向地质剖面图Q4edl-全新世风成堆积,Qp1m-早更新世汨罗组,E-古近纪。Figure 5. NS-trending geological profile of the bedrock surface of Chishan Uplift and field photos of Miluo FormationQ4edl-wind accumulation of Holocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.
图 5 赤山凸起基岩面海拔75 m左右南北向地质剖面图Q4edl-全新世风成堆积,Qp1m-早更新世汨罗组,E-古近纪。Figure 5. NS-trending geological profile of the bedrock surface of Chishan Uplift and field photos of Miluo FormationQ4edl-wind accumulation of Holocene,Qp1m-Miluo Formation of Early Pleistocene,E-Paleogene.①层为粉砂质黏土,局部含泥质粉砂岩细碎石,厚度变化较大,约为0.4~1.2 m,结构松散,明显为第四纪全新统地层,风成沉积。
②层为泥砾层,属泥、砾混杂堆积,成分复杂,沉积散乱不规律,可能为坡积与冲积堆积混合地层,结构较松散,厚度为0.2~0.6 m,为过渡层。
③层为网纹红土,成分为粉砂质黏土,网纹发育,明显经过湿热化,多呈水平状,并见黄色斑点和铁质结核,厚度约为2.0 m,并与下部④层表部的含砾网纹红土呈过渡关系,河湖相,为早更新统汨罗组冲湖积堆积。
④层为灰红色砾石层,砾石成分以脉石英(70%左右)为主,其次为砂岩、硅质岩,含量高达85%左右,粒径一般1~3 cm,形状以次圆—圆状为主,少量次棱角状,分选较好,砾石多为等轴或近等轴状,定向性不明显,砾石间孔隙充填物多为细—粗砂,河湖相,为早更新统汨罗组冲湖积堆积。厚度约为10~20 m。
⑤层为红褐色泥质粉砂岩,为古近纪红层,未揭露层底。
2.4 分析与讨论
根据第四纪古气候有关研究[14, 21],早更新世早期和晚期气候皆是先干冷后暖湿;中更新世早期为冷干,中期为暖湿,晚期为湿热;晚更新世为干冷、严寒;全新世早期为干冷向暖湿过渡。干冷气候,雨水少,水位低,湖泊面积小,湖区大部分裸露成陆地,处于剥蚀状态,沉积以河流相和河湖相为主;暖湿气候,降雨量多,汇入湖区径流多,湖水位高,湖面广;湿热气候,温度高,蒸发强烈,湿度大,降雨和汇入湖区径流相比暖湿气候明显偏少,湖水位也较低,且地表土层在长期湿热气候作用下易发生网纹化。
结合第四纪古气候、钻孔揭露的地层资料和地质调查测绘,赤山凸起不同高程区沉积地层差异可解释为:
(1)由于早更新世早期、晚期古气候皆为先干冷后暖湿,若赤山凸起未发生隆起或沉陷,在两个暖湿气候时段,降雨充沛,湖区水位较高,赤山部分或全部区域应处于水下接受早更新世早期和晚期河湖相地层沉积,但本次钻孔揭露发现,赤山只沉积了早更新世晚期汨罗组地层,未见早期地层。可能是早更新世早期赤山凸起地势较高,早期一直裸露于湖水面,处于剥蚀状态,而未沉积早期地层;早更新世早期末至晚期早段,赤山凸起发生了沉陷,晚期暖湿气候来临后,湖区水位上涨,整个赤山凸起绝大部分区域淹没在湖水中,开始接受晚期汨罗组地层,且由于降雨量大,流速快,其沉积的地层多为粒径相对较粗的砂卵砾石,且呈现出岛内低地势区域沉积砾石层厚、高地势区域薄的特征;早更新世末期气候慢慢向干冷气候过渡,湖水位开始下降,水流搬运能力变弱,赤山凸起沉积的汨罗组地层颗粒变细,赤山高地势区域露出水面不再接受汨罗组地层沉积,导致岛内高地势汨罗组地层较薄而低地势相对较厚,这与本次揭露的3个高程区汨罗组沉积颗粒特性下部粗上部细及厚度变化特征是相一致的。
(2)中更新世早期,气候干冷,径流小,入湖水流挟沙能力弱、颗粒粒径小,湖区水位较低,湖面窄,赤山凸起发生小幅沉陷后稳定,使得岛内基岩面海拔55 m左右及以下区域淹没在水中,开始接受中更新统新开铺组地层,到中更新世早期晚段赤山强烈凸起,虽然气候慢慢进入暖湿,降雨增多、入湖径流大,湖区水位上升,但赤山凸起地势较高区域裸露地表。这与赤山凸起中沉积的新开铺组地层为细粒黏性土且厚度较薄较为一致。
(3)中更新世中期,赤山构造稳定,气候暖湿,湖区水位上升,但由于赤山凸起地势较高,仅地势相对较低区域(主要为基岩面海拔55 m以下区域,以上区域是否在内无法确定)再次淹没在水中,沉积白沙井组地层,其他高地势区域处于剥蚀状态,但由于目平湖凹陷的持续扩大,挟沙水流进入湖区后流速变慢,而无法将大颗粒的砾石等搬运至赤山区域,导致该时期内沉积的地层颗粒粒径较小。
(4)中更新世晚期气候转为湿热,湖区水位稍有下降,导致基岩面海拔0 m左右及以上区域皆已裸露水表成陆,处于剥蚀状态,导致赤山凸起地表裸露的汨罗组、新开铺组和白沙井组地层厚度变薄,甚至部分区域部分时代地层被完全剥蚀;同时受湿热气候长期作用,赤山裸露区域表部土层发生网纹化。与本次钻探揭露更新世最顶部地层皆存在一定厚度的网纹化土层相一致。
3. 文化遗址
3.1 文化遗址特征
据湖南省文物考古研究所联合益阳市文物处等单位对赤山凸起旧石器遗址长期考古调查研究[22],赤山凸起共发现旧石器遗址10余处,大部分发现于地表,少部分深埋于第四纪地层中,且主要位于地势相对较低区域的第四纪中更新世白沙井组或新开铺组上覆堆积层中。
岛内比较有代表性的两个旧石器遗址枫树嘴遗址和杨腊丘遗址,两地点南北相距约2 km。揭露的文化层剖面厚4~5 m,地层堆积基本相同,自上而下分别为(图6):
![]() 图 6 赤山凸起旧石器遗址地质剖面图Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,E-古近纪。Figure 6. Geological profile of paleolithic sites in Chishan UpliftQ4edl- Holocene aeolian deposits,Qp2b-Baishajing Formation of Middle Pleistocene,E-Paleogene.
图 6 赤山凸起旧石器遗址地质剖面图Q4edl-全新世风成堆积,Qp2b-中更新世白沙井组,E-古近纪。Figure 6. Geological profile of paleolithic sites in Chishan UpliftQ4edl- Holocene aeolian deposits,Qp2b-Baishajing Formation of Middle Pleistocene,E-Paleogene.第①层,灰褐色粉砂黏土,土质疏松,含有少量青花、白釉、酱釉等瓷器碎片,属近现代表土层,厚0.4~0.5 m;
第②层,浅红色黏土粉砂,土质较疏松,厚0.3~0.8 m,出土有石制品,属上文化层;
第③层,黄—红褐色黏土粉砂,较细的灰黄色网纹较密集分布其中,含大量褐色的铁锰结核颗粒,土质较紧密,厚0.6~1.5 m,出土有较多石制品,属主要文化层;
第④层,网纹红土,呈褐红色,分布其中的灰黄色网纹较上层粗、稀疏,铁锰结核颗粒渐少,但与上部层位无明显界限,呈渐变状态,土质紧密,厚1.0~1.5 m,出土有石制品;
第⑤层,网纹红土,呈赭红色,灰白色网纹较上部粗,极少或不含铁锰结核,土质紧密,基本不见石制品,已发掘0.4~0.5 m,其下约5 m至底部含卵砂质黏土。
石制品主要出自第③层和第④层上部,第②层中有少量,其余层位没有遗物发现。可见较多石制品埋藏在均质红土层至铁锰结核弱网纹红土层,也有部分石制品出自网纹红土层的上部,一些采集的石器表面清晰见有网状印迹。从出土层位分析,遗址可能有两个不同的人类活动时期,但含铁锰结核颗粒较多的第③层网纹红土堆积所对应的阶段应是赤山凸起区域古人类活动最为频繁的时间段。因此,赤山凸起内旧石器遗址文化层主要位于网纹红土的上部及上覆堆积土层中,遗址的年代应较网纹红土(即中更新世白沙井组二元结构上部黏性土层)形成的时代偏晚,属中更新世晚期至晚更新世早期,绝对年代大致为200~100 ka,在考古学上大致与旧石器时代早期晚段或旧石器中期相当。
赤山凸起区域并未发现旧石器晚期和新石器遗址,即晚更新世早期之后至新石器时代结束,赤山凸起区域皆无人类活动。但湖区其他地区却发现众多新石器遗址[23-24],据不完全统计,遗址约有130多处。可能是晚更新世早期,世界性低海面出现,引发长江干流河床发生溯源侵蚀,导致洞庭湖出口河床发生强烈下切,洞庭湖水位剧降,湖水被迅速排干,湖面急剧萎缩,湖盆底部大多露出成为陆地,形成一片由滨湖阶地环绕的河网平原[25-26],人类天性近水生存,从而搬离海拔较高、离水源较远的赤山凸起,而居住在离水较近的河网平原,也就造成赤山凸起无新石器遗址而湖区地势低矮河网平原却广泛分布。
3.2 分析与讨论
赤山凸起旧石器文化遗址发现于第③层和第④层上部,第②层只有少量,其余层位没有发现,表明:
(1)中更新世晚期初,赤山凸起基岩面海拔0 m左右及以上区域已裸露湖面,并开始有人类活动,由于古人类临水而居的天性,其活动主要集中在岛内地势相对较低的临水区域,使得刚刚凸起露出地表的中更新世中期白沙井组地层顶部土层中存在人类活动的痕迹,并遗留下旧石器遗址,表现为第④层上部存在旧石器时代石制品。
(2)旧石器遗址文化层在第④层上部、第③层和第②层土中皆有,可见赤山凸起从中更新世晚期至第②层土沉积(全新世早期)过程中,一直存在人类活动,即这一时期岛内基岩面海拔0 m及以上区域一直裸露水表,处于剥蚀状态,且中更新世晚期气候湿热,晚更新世干冷、严寒,全新世早期由干冷缓慢过渡至暖湿,湖区这一时期水位较低,特别是晚更新世洞庭湖已演变成河网平原,同时该时期内赤山凸起构造活动较平静[5],其被湖水淹没可能性极小,这一时期内沉积的地层应主要为风成堆积。
(3)赤山凸起基岩面0 m及以上区域第四系地层顶部存在一定厚度风成堆积土,应大部分来源于岛内相对较高的第四系沉积和岛内丘陵风化剥蚀;高海拔区域的汨罗组和新开铺组上部部分地层被风化剥蚀,甚至部分地层分布局限且局部残留,厚度也较薄,顶部一般存在一定厚度的风化坡积产物。
4. 结论
(1)赤山凸起第四纪沉积除了浅表层存在较薄的全新统外,其他皆为更新世地层。其中基岩面海拔75 m左右区域沉积有早更新世晚期汨罗组地层,缺失中、晚更新世地层;55 m左右区域沉积有早更新世晚期汨罗组和中更新世早期新开铺组地层,缺失中更新世中期、晚期及晚更新世地层;0 m左右区域沉积有早更新世汨罗组和中更新世早期开铺组、中期白沙井组地层,缺失晚更新世地层;且不同基岩面海拔区相互之间组成镶嵌阶地。
(2)赤山凸起早更新世晚期构造活动表现为先凹陷后稳定,中更新世早期—中期末表现为稳定—凹陷—稳定—凸起—稳定。
(3)中更新世晚期初,赤山凸起基岩面海拔0 m左右区域绝大部分已裸露成陆,人类活动开始,并在这些区域地层顶部及之后沉积地层中遗留下连续性较好的旧石器遗址,即中更新世晚期至全新世早期赤山凸起均处于剥蚀状态,凸起内中更新世晚期及之后沉积的第四纪地层应主要为风成堆积。
本文通过多角度分析论证,揭示了赤山凸起第四纪以来构造沉积演变特性,但仍有一些疑问之处,如早更新世晚期赤山凸起凹陷下沉持续时间、基岩面海拔75及55 m左右区域是否曾经沉积过中更新世早期和中期地层(只是后来被完全剥蚀)等,需加强相关论证研究。
-
![]()
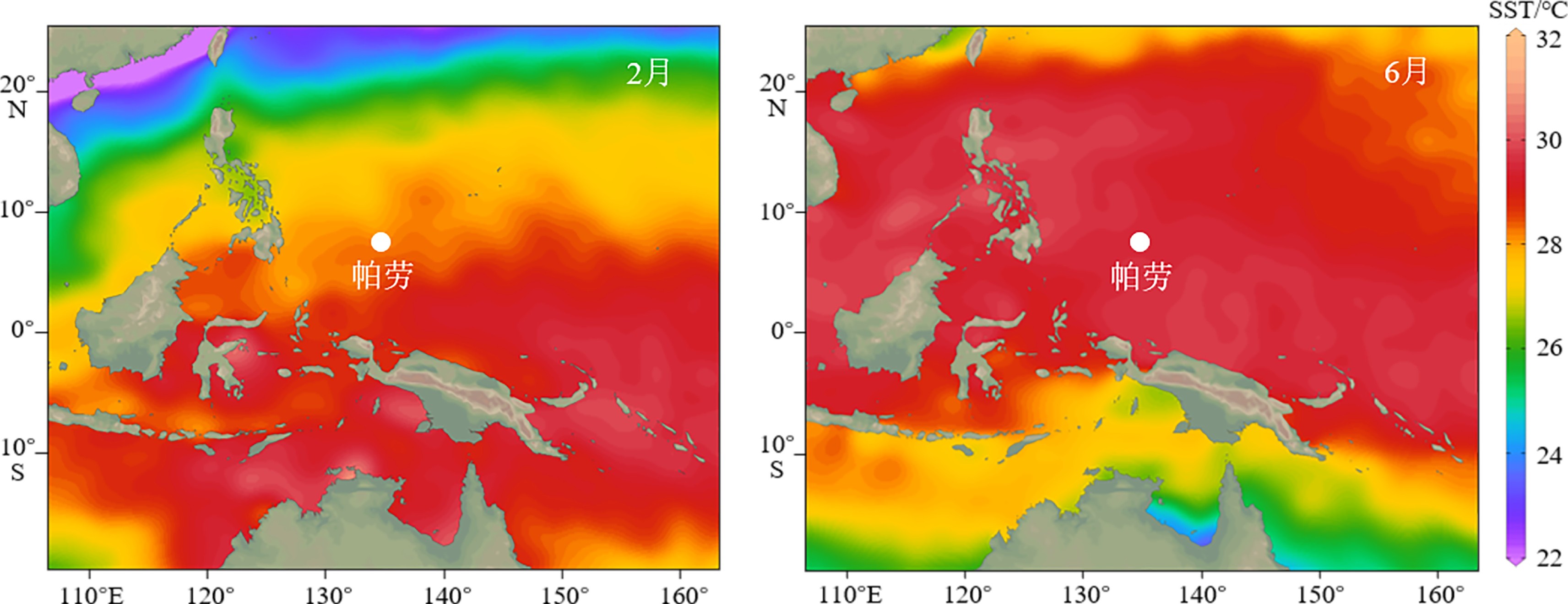
图 1 西北太平洋2月(最冷月)与6月(最暖月)平均SST分布(1955—2017年)和帕劳位置示意图
图中白色标记点为帕劳,SST数据来自WOA:http://odv.awi.de/data/ocean/
Figure 1. The monthly average SST of February (coldest month) and June (warmest month) in the northwestern Pacific (1955—2017) and the location of Palau
The location of the Palau is marked by white point , SST data is obtained from WOA: http://odv.awi.de/data/ocean/
![]()
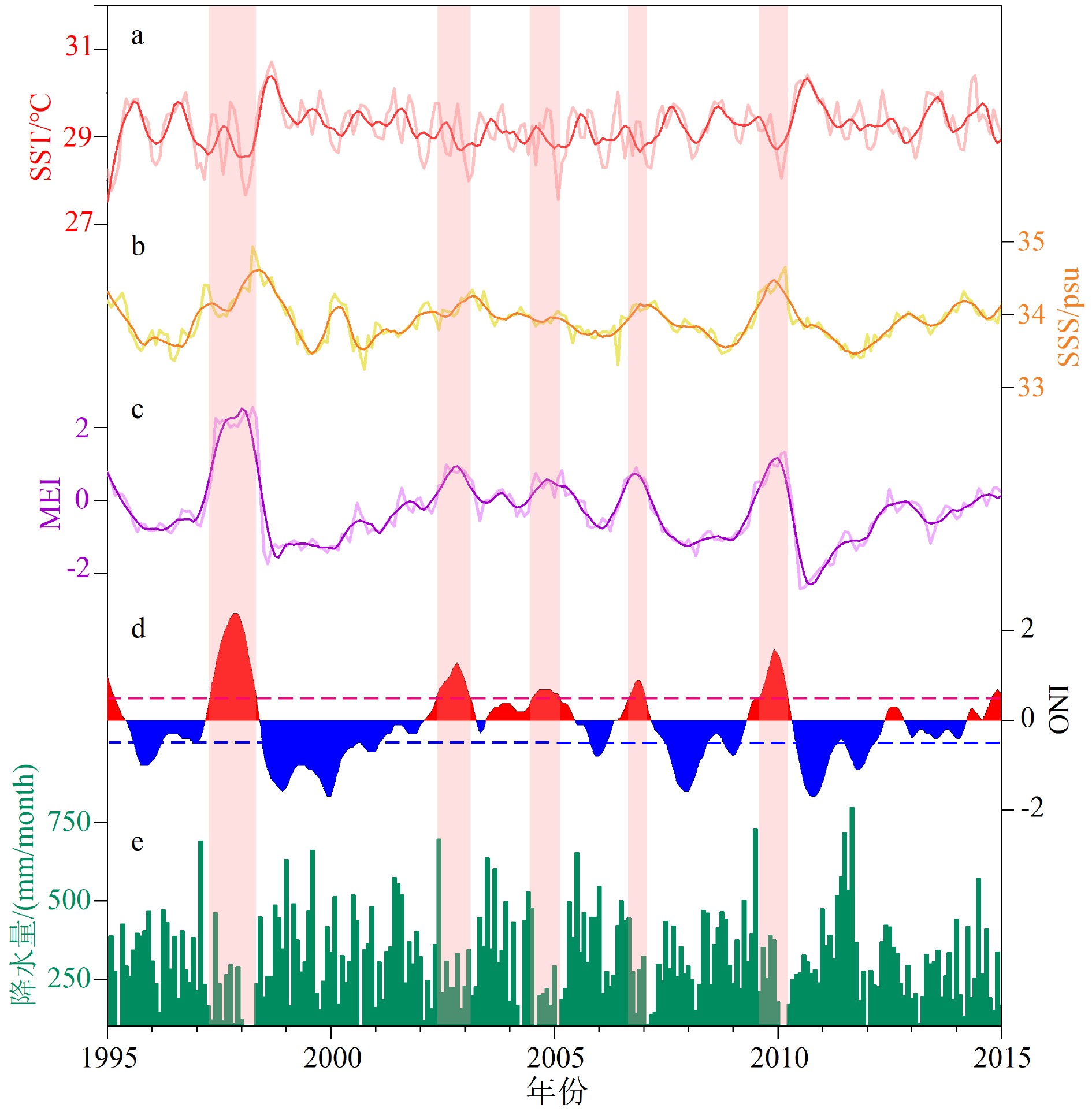
图 2 帕劳器测资料和ENSO指数的对比(1995—2015年)
a. 帕劳月平均SST记录,b. 帕劳月平均SSS记录,c. MEI记录,d. ONI记录,e. 帕劳月平均降水记录;图中a、b、c中的粗实线均为12点滑动平均曲线,图d中红色和蓝色虚线分别为El Niño事件(0.5 ℃)和La Niña事件(−0.5 ℃)的检测阈值,淡红色阴影表示El Niño事件。
Figure 2. The comparison of instrumental data of Palau with indices of ENSO (1995—2015)
a. monthly average SST of Palau, b. monthly average SSS of Palau, c. MEI, d. ONI, e. monthly average Precipitation of Palau. The thick lines in Fig.2a, b and c are the 12-point moving average curves; The red and blue dotted lines in the Fig.2d mark the threshold value for El Niño event (0.5 ℃) and La Niña event (−0.5 ℃), respectively. The light red shading represents El Niño event.
![]()
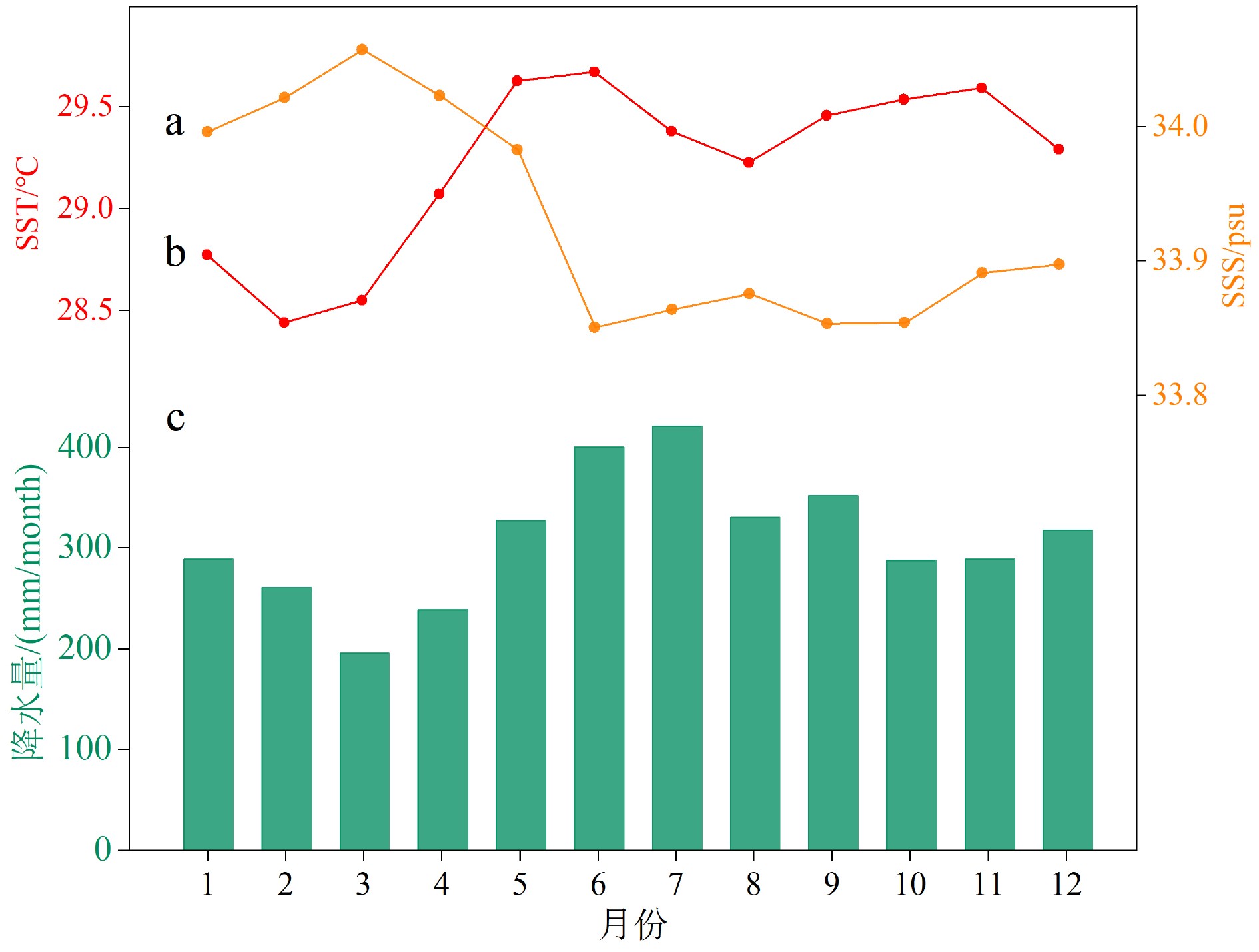
图 3 1995—2015年帕劳多年月平均SSS(a)、SST(b)和降水(c)
Figure 3. The multi-a monthly average SSS (a), SST (b) and Precipitation (c) of Palau (1995—2015)
![]()
图 4 帕劳砗磲PL-1壳体(a)、内层薄片(b)及内层激光共聚焦图像(c)
Figure 4. Photograph of T. gigas PL-1(a), slab of PL-1(b), and laser scanning confocal image of inner shell(c)
![]()
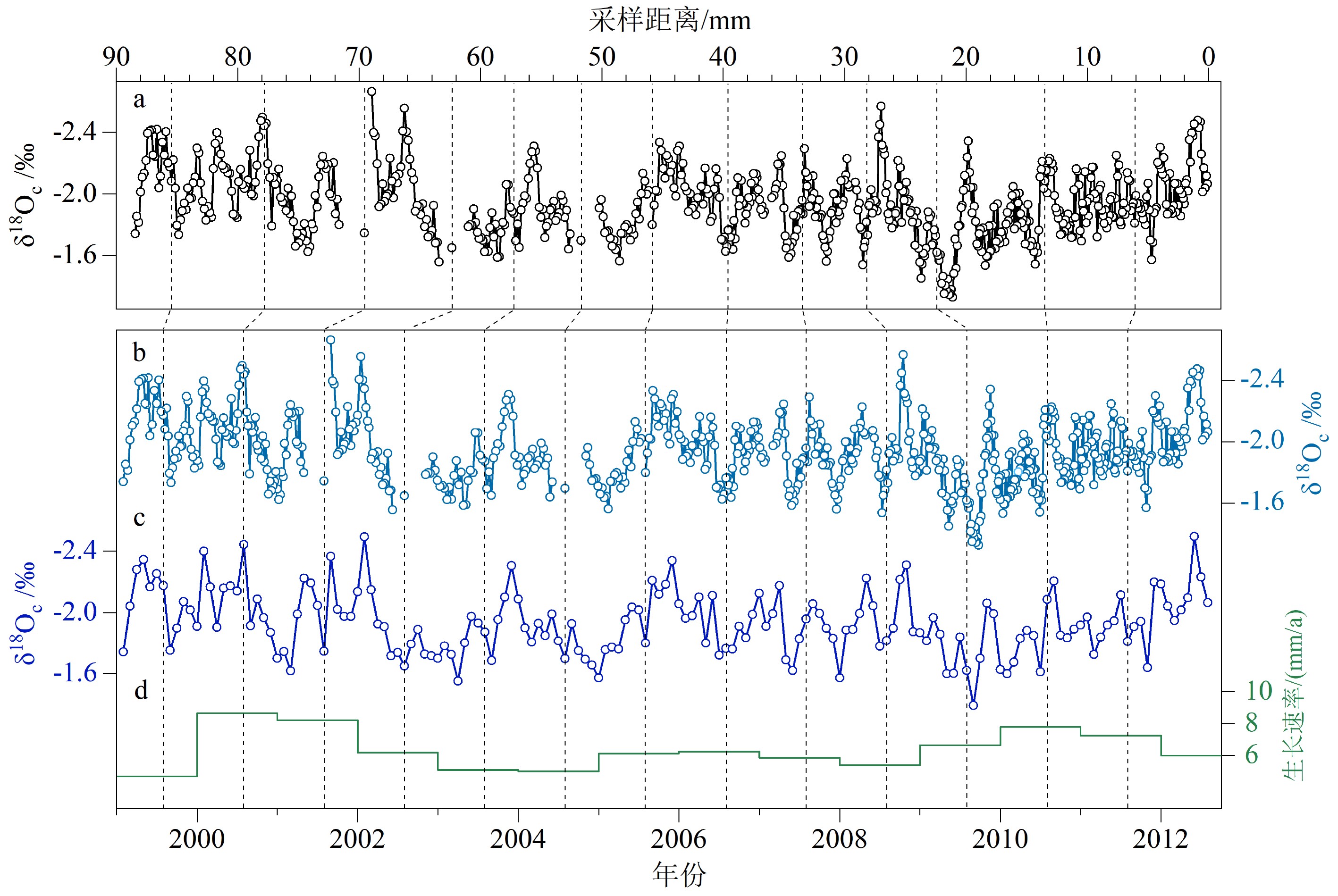
图 5 PL-1内层δ18Oc(a)、δ18Oc年龄模型(b)、12点插值的δ18Oc(c)、内层壳体的年生长速率(d)
Figure 5. The δ18Oc profile (a), the chronology of δ18Oc profile (b), the 12-point profile of δ18Oc (c) and the annual growth rate of inner shell of PL-1 (d)
![]()
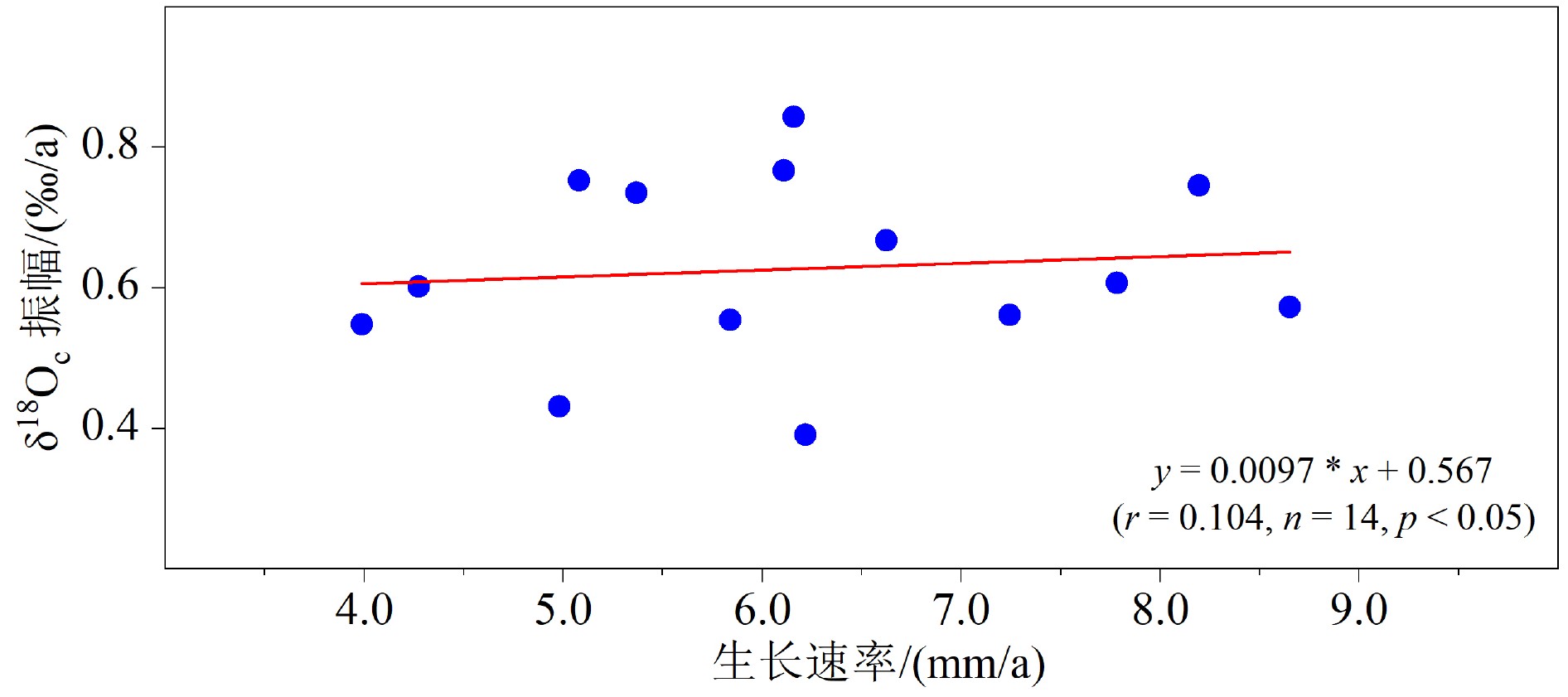
图 6 PL-1 δ18Oc年振幅和年生长速率的相关性
Figure 6. The correlation between the δ18Oc annual amplitude and annual growth rate of PL-1
![]()
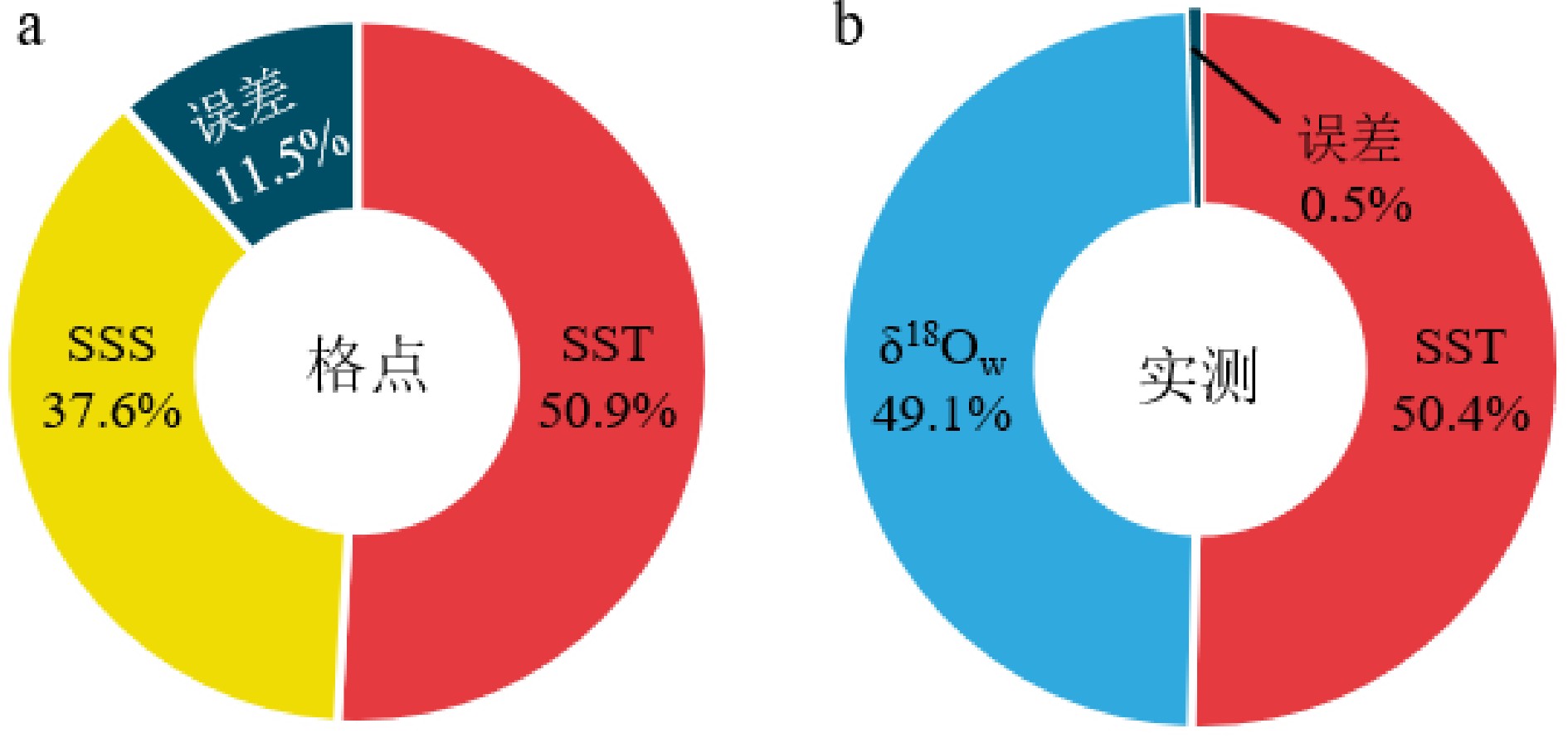
图 7 格点SST和SODA SSS(a)、实测SST和δ18Ow(b)对δ18Oc年振幅的贡献
Figure 7. The contributions of instrumental SST and SODA SSS (a), suit SST and suit δ18Ow (b) to the annual amplitude of δ18Oc
![]()
图 8 12点δ18Oc与SST(a),SSS(b),降水(c)之间的对比
蓝色实线为12点插值的δ18Oc,红色实线为SST,黄色实线为SSS,绿色实线为降水。
Figure 8. The comparison of 12-point profile of δ18Oc with SST (a), SSS (b), and Precipitation (c)
The blue solid line is the 12-point profile of δ18Oc, the red solid line is SST, the yellow solid line is SSS, the green solid line is Precipitation.
![]()
图 9 δ18OA(a)与SSTA(b)、降水异常(c)、SSSA(d)、MEI(e)、SOI(f)的对比
图中粗实线均为1年滑动平均曲线;蓝色和红色阴影分别代表由ONI指示的La Niña与El Niño事件,带斜杠的蓝色阴影代表δ18OA未检测出的La Niña;δ18OA、SSSA、MEI的纵轴为逆序坐标。
Figure 9. The comparison of δ18OA (a) with SSTA (b), Precipitation anomaly (c), SSSA (d), MEI (e), SOI (f)
All of the thick solid lines in the figure are the 1-a moving average curves; the blue and red shadings represent the La Niña and El Niño events indicated by ONI, respectively; the blue bar with slashes represents the La Niña event that is not detected by δ18OA; the vertical axes of δ18OA, SSSA and MEI are reversed.
-
[1] Andreasson F P, Schmitz B. Temperature seasonality in the early middle Eocene North Atlantic region: Evidence from stable isotope profiles of marine gastropod shells [J]. GSA Bulletin, 2000, 112(4): 628-640. doi: 10.1130/0016-7606(2000)112<628:TSITEM>2.0.CO;2
[2] Yan H, Sun L G, Shao D, et al. Seawater temperature seasonality in the South China Sea during the late Holocene derived from high-resolution Sr/Ca ratios of Tridacna gigas [J]. Quaternary Research, 2015, 83(2): 298-306. doi: 10.1016/j.yqres.2014.12.001
[3] 林而达, 许吟隆, 蒋金荷, 等. 气候变化国家评估报告(Ⅱ): 气候变化的影响与适应[J]. 气候变化研究进展, 2006, 2(2):51-56 doi: 10.3969/j.issn.1673-1719.2006.02.001 LIN Erda, XU Yinlong, JIANG Jinhe, et al. National Assessment Report of Climate Change (Ⅱ): Climate change impacts and adaptation [J]. Advances in Climate Change Research, 2006, 2(2): 51-56. doi: 10.3969/j.issn.1673-1719.2006.02.001
[4] Collins M, An S I, Cai W J, et al. The impact of global warming on the tropical Pacific Ocean and El Niño [J]. Nature Geoscience, 2010, 3(6): 391-397. doi: 10.1038/ngeo868
[5] Guo Y P, Tan Z M. The Hadley circulation regime change: Combined effect of the Western Pacific warming and increased ENSO amplitude [J]. Journal of Climate, 2018, 31(23): 9739-9751. doi: 10.1175/JCLI-D-18-0306.1
[6] Hongo C, Kurihara H, Golbuu Y. Coral boulders on Melekeok reef in the Palau Islands: An indicator of wave activity associated with tropical cyclones [J]. Marine Geology, 2018, 399: 14-22. doi: 10.1016/j.margeo.2018.02.004
[7] Schöne B R, Castro A D F, Fiebig J, et al. Sea surface water temperatures over the period 1884-1983 reconstructed from oxygen isotope ratios of a bivalve mollusk shell (Arctica islandica, southern North Sea) [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2004, 212(3-4): 215-232. doi: 10.1016/j.palaeo.2004.05.024
[8] Patterson W P, Dietrich K A, Holmden C, et al. Two millennia of North Atlantic seasonality and implications for Norse colonies [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(12): 5306-5310. doi: 10.1073/pnas.0902522107
[9] Yamanashi J, Takayanagi H, Isaji A, et al. Carbon and oxygen isotope records from Tridacna derasa shells: toward establishing a reliable proxy for sea surface environments [J]. PLoS ONE, 2016, 11(6): e0157659. doi: 10.1371/journal.pone.0157659
[10] Xu J, Kuhnt W, Holbourn A, et al. Indo-Pacific warm pool variability during the Holocene and Last Glacial Maximum [J]. Paleoceanography, 2010, 25(4): PA4230.
[11] 孙有斌, 郭飞. 中国黄土记录的季风快速变化[J]. 第四纪研究, 2017, 37(5):963-973 doi: 10.11928/j.issn.1001-7410.2017.05.04 SUN Youbin, GUO Fei. Rapid monsoon changes recorded by Chinese loess deposits [J]. Quaternary Sciences, 2017, 37(5): 963-973. doi: 10.11928/j.issn.1001-7410.2017.05.04
[12] Greenland Ice-core Project (GRIP) Members. Climate instability during the last interglacial period recorded in the GRIP ice core [J]. Nature, 1993, 364(6434): 203-207. doi: 10.1038/364203a0
[13] Neukom R, Steiger N, Gómez-Navarro J J, et al. No evidence for globally coherent warm and cold periods over the preindustrial Common Era [J]. Nature, 2019, 571(7766): 550-554. doi: 10.1038/s41586-019-1401-2
[14] Jones D S, Williams D F, Romanek C S. Life history of symbiont-bearing giant clams from stable isotope profiles [J]. Science, 1986, 231(4733): 46-48. doi: 10.1126/science.231.4733.46
[15] Rosewater J. The Family Tridacnidae in the Indo-Pacific[M]. The Philippines: Department of Mollusks, Academy of Natural Sciences of Philadelphia, 1965: 374-396.
[16] Jameson S C. Early life history of the giant clams Tridacna crocea Lamarck, Tridacna maxima (Roding), and Hippopus hippopus (Linnaeus) [J]. Pacific Science, 1976, 30(3): 219-233.
[17] Aharon P, Chappell J, Compston W. Stable isotope and sea-level data from New Guinea supports Antarctic ice-surge theory of ice ages [J]. Nature, 1980, 283(5748): 649-651. doi: 10.1038/283649a0
[18] Driscoll R E. PaleoENSO reconstructions of the Holocene and Last Glacial Period[D]. Doctor Dissertation of University of Edinburgh, 2015.
[19] Gannon M E, Pérez-Huerta A, Aharon P, et al. A biomineralization study of the Indo-Pacific giant clam Tridacna gigas [J]. Coral Reefs, 2017, 36(2): 503-517. doi: 10.1007/s00338-016-1538-5
[20] 晏宏, 刘成程. 砗磲地球化学与古气候学研究进展[J]. 第四纪研究, 2017, 37(5):1077-1090 doi: 10.11928/j.issn.1001-7410.2017.05.15 YAN Hong, LIU Chengcheng. Review on Tridacna geochemistry and paleoclimate research [J]. Quaternary Sciences, 2017, 37(5): 1077-1090. doi: 10.11928/j.issn.1001-7410.2017.05.15
[21] 梅衍俊, 邵达, 刘文齐, 等. 南海砗磲壳体成分及生物有机特征分析[J]. 中国科学技术大学学报, 2018, 48(7):550-559 doi: 10.3969/j.issn.0253-2778.2018.07.005 MEI Yanjun, SHAO Da, LIU Wenqi, et al. Analysis of the components and biological organic characteristics of Tridacna spp. shells from South China Sea [J]. Journal of University of Science and Technology of China, 2018, 48(7): 550-559. doi: 10.3969/j.issn.0253-2778.2018.07.005
[22] 晏宏, 邵达, 王玉宏, 等. 南海西沙大砗磲高分辨率Sr/Ca温度计及其意义[J]. 地球环境学报, 2011, 2(2):381-386 YAN Hong, SHAO Da, WANG Yuhong, et al. High resolution Sr/Ca profile of Tridacna gigas from Xisha Islands of South China Sea and its potential application on sea surface temperature reconstruction [J]. Journal of Earth Environment, 2011, 2(2): 381-386.
[23] Aharon P, Chappell J. Carbon and oxygen isotope probes of reef environment histories[M]//Barnes D J. Perspectives on Coral Reefs. Townsville, Australia: Brian Clouston, 1983: 1-15.
[24] Yoshimura T, Tamenori Y, Suzuki A, et al. Element profile and chemical environment of sulfur in a giant clam shell: Insights from μ-XRF and X-ray absorption near-edge structure [J]. Chemical Geology, 2013, 352: 170-175. doi: 10.1016/j.chemgeo.2013.05.035
[25] Aubert A, Lazareth C E, Cabioch G, et al. The tropical giant clam Hippopus hippopus shell, a new archive of environmental conditions as revealed by sclerochronological and δ18O profiles [J]. Coral Reefs, 2009, 28(4): 989-998. doi: 10.1007/s00338-009-0538-0
[26] Warter V, Erez J, Müller W. Environmental and physiological controls on daily trace element incorporation in Tridacna crocea from combined laboratory culturing and ultra-high resolution LA-ICP-MS analysis [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 496: 32-47. doi: 10.1016/j.palaeo.2017.12.038
[27] Watanabe T, Oba T. Daily reconstruction of water temperature from oxygen isotopic ratios of a modern Tridacna shell using a freezing microtome sampling technique [J]. Journal of Geophysical Research: Oceans, 1999, 104(C9): 20667-20674. doi: 10.1029/1999JC900097
[28] Duprey N, Lazareth C E, Dupouy C, et al. Calibration of seawater temperature and δ18Oseawater signals in Tridacna maxima’s δ18Oshell record based on in situ data [J]. Coral Reefs, 2015, 34(2): 437-450. doi: 10.1007/s00338-014-1245-z
[29] Komagoe T, Watanabe T, Shirai K, et al. Geochemical and microstructural signals in giant clam Tridacna maxima recorded typhoon events at Okinotori Island, Japan [J]. Journal of Geophysical Research: Biogeosciences, 2018, 123(5): 1460-1474. doi: 10.1029/2017JG004082
[30] Bayer S, Beierlein L, Morán G A, et al. Late Quaternary climatic variability in northern Patagonia, Argentina, based on δ18O of modern and fossil shells of Amiantis purpurata (Bivalvia, Veneridae) [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 560: 110012. doi: 10.1016/j.palaeo.2020.110012
[31] Watanabe T, Suzuki A, Kawahata H, et al. A 60-year isotopic record from a mid-Holocene fossil giant clam (Tridacna gigas) in the Ryukyu Islands: physiological and paleoclimatic implications [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2004, 212(3-4): 343-354. doi: 10.1016/S0031-0182(04)00358-X
[32] Ayling B F, Chappell J, Gagan M K, et al. ENSO variability during MIS 11 (424-374 ka) from Tridacna gigas at Huon Peninsula, Papua New Guinea [J]. Earth and Planetary Science Letters, 2015, 431: 236-246. doi: 10.1016/j.jpgl.2015.09.037
[33] Yan H, Shao D, Wang Y H, et al. Sr/Ca profile of long-lived Tridacna gigas bivalves from South China Sea: A new high-resolution SST proxy [J]. Geochimica et Cosmochimica Acta, 2013, 112: 52-65. doi: 10.1016/j.gca.2013.03.007
[34] Hori M, Sano Y, Ishida A, et al. Middle Holocene daily light cycle reconstructed from the strontium/calcium ratios of a fossil giant clam shell [J]. Scientific Reports, 2015, 5: 8734. doi: 10.1038/srep08734
[35] Yan H, Liu C C, An Z S, et al. Extreme weather events recorded by daily to hourly resolution biogeochemical proxies of marine giant clam shells [J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(13): 7038-7043. doi: 10.1073/pnas.1916784117
[36] Schwartzmann C, Durrieu G, Sow M, et al. In situ giant clam growth rate behavior in relation to temperature: A one-year coupled study of high-frequency noninvasive valvometry and sclerochronology [J]. Limnology and Oceanography, 2011, 56(5): 1940-1951. doi: 10.4319/lo.2011.56.5.1940
[37] Aharon P, Chappell J. Oxygen isotopes, sea level changes and the temperature history of a coral reef environment in New Guinea over the last 105 years [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1986, 56(3-4): 337-379. doi: 10.1016/0031-0182(86)90101-X
[38] Grossman E L, Ku T L. Oxygen and carbon isotope fractionation in biogenic aragonite: temperature effects [J]. Chemical Geology, 1986, 59: 59-74. doi: 10.1016/0168-9622(86)90057-6
[39] Welsh K, Elliot M, Tudhope A, et al. Giant bivalves (Tridacna gigas) as recorders of ENSO variability [J]. Earth and Planetary Science Letters, 2011, 307(3-4): 266-270. doi: 10.1016/j.jpgl.2011.05.032
[40] Arias-Ruiz C, Elliot M, Bézos A, et al. Geochemical fingerprints of climate variation and the extreme La Niña 2010-11 as recorded in a Tridacna squamosa shell from Sulawesi, Indonesia [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 487: 216-228. doi: 10.1016/j.palaeo.2017.08.037
[41] Duprey N, Galipaud J C, Cabioch G, et al. Isotopic records from archeological giant clams reveal a variable climate during the southwestern Pacific colonization ca. 3.0 ka BP [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 404: 97-108. doi: 10.1016/j.palaeo.2014.04.002
[42] Yan H, Liu C C, Zhang W C, et al. ENSO variability around 2000 years ago recorded by Tridacna gigas δ18O from the South China Sea [J]. Quaternary International, 2017, 452: 148-154. doi: 10.1016/j.quaint.2016.05.011
[43] Hu Y, Sun X M, Cheng H, et al. Evidence from giant-clam δ18O of intense El Ninõ-Southern Oscillation-related variability but reduced frequency 3700 years ago [J]. Climate of the Past, 2020, 16(2): 597-610. doi: 10.5194/cp-16-597-2020
[44] Conroy J L, Noone D, Cobb K M, et al. Paired stable isotopologues in precipitation and vapor: A case study of the amount effect within western tropical Pacific storms [J]. Journal of Geophysical Research: Atmospheres, 2016, 121(7): 3290-3303. doi: 10.1002/2015JD023844
[45] Dansgaard W. Stable isotopes in precipitation [J]. Tellus, 1964, 16(4): 436-468. doi: 10.3402/tellusa.v16i4.8993
[46] Ma X L, Yan H, Fei H B, et al. A high-resolution δ18O record of modern Tridacna gigas bivalve and its paleoenvironmental implications [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 554: 109800. doi: 10.1016/j.palaeo.2020.109800
[47] Colin P L. Ocean Warming and the Reefs of Palau [J]. Oceanography, 2018, 31(2): 126-135.
[48] Bruno J, Siddon C, Witman J, et al. El Niño related coral bleaching in Palau, Western Caroline Islands [J]. Coral Reefs, 2001, 20(2): 127-136. doi: 10.1007/s003380100151
[49] Martin L E, Dawson M N, Bell L J, et al. Marine lake ecosystem dynamics illustrate ENSO variation in the tropical western Pacific [J]. Biology Letters, 2006, 2(1): 144-147. doi: 10.1098/rsbl.2005.0382
[50] Colin P L. Marine Environments of Palau[M]. San Diego: Indo-Pacific Press, 2009: 15-25.
[51] Golbuu Y, Bauman A, Kuartei J, et al. The state of coral reef ecosystems of Palau [J]. The state of coral reef ecosystems of the United States and Pacific freely associated states, 2005, 2005: 488-507.
[52] Hardy J T, Hardy S A. Ecology of Tridacna in Palau [J]. Pacific Science, 1969, XXIII: 467-472.
[53] Colin P L. Marine Environments of Palau[M]. San Diego: Indo-Pacific Press, 2009: 365-366.
[54] Grottoli A G. Monthly resolved stable oxygen isotope record in a Palauan sclerosponge Acanthocheatetes wellsi for the period of 1977-2001[C]//Proceedings of the 10th International Coral Reef Symposium. Okinawa, Japan, 2006: 572-579.
[55] Iijima H, Kayanne H, Morimoto M, et al. Interannual sea surface salinity changes in the western Pacific from 1954 to 2000 based on coral isotope analysis [J]. Geophysical Research Letters, 2005, 32(4): L04608.
[56] Osborne M C, Dunbar R B, Mucciarone D A, et al. Regional calibration of coral-based climate reconstructions from Palau, West Pacific Warm Pool (WPWP) [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 386: 308-320. doi: 10.1016/j.palaeo.2013.06.001
[57] Wu H C, Grottoli A G. Stable oxygen isotope records of corals and a sclerosponge in the Western Pacific warm pool [J]. Coral Reefs, 2010, 29(2): 413-418. doi: 10.1007/s00338-009-0576-7
[58] Osborne M C, Dunbar R B, Mucciarone D A, et al. A 215-yr coral δ18O time series from Palau records dynamics of the West Pacific Warm Pool following the end of the Little Ice Age [J]. Coral Reefs, 2014, 33(3): 719-731. doi: 10.1007/s00338-014-1146-1
[59] Grottoli A G, Adkins J F, Panero W R, et al. Growth rates, stable oxygen isotopes (δ18O), and strontium (Sr/Ca) composition in two species of Pacific sclerosponges (Acanthocheatetes wellsi and Astrosclera willeyana) with δ18O calibration and application to paleoceanography [J]. Journal of Geophysical Research: Oceans, 2010, 115(C6): C06008.
[60] Pätzold J, Heinrichs J P, Wolschendorf K, et al. Correlation of stable oxygen isotope temperature record with light attenuation profiles in reef-dwelling Tridacna shells [J]. Coral Reefs, 1991, 10(2): 65-69. doi: 10.1007/BF00571825
[61] Jew N P, Dodrill T, Fitzpatrick S M. Evaluating the efficacy of the mollusc Tridacna crocea for reconstructing ancient sea-surface temperatures in the Rock Islands of Palau, Micronesia [J]. Archaeology in Oceania, 2019, 54(2): 107-119. doi: 10.1002/arco.5182
[62] Dodrill T N, Lassuy M G, Jew N P, et al. Stable Oxygen Isotope (δ18O) Analyses and Paleoenvironmental Reconstructions from Mollusks in Palau, Micronesia[C]//Proceedings of the 81st Annual Meeting of the Society for American Archaeology. Orlando, FL, 2016.
[63] Maes C. Salinity variability in the equatorial Pacific Ocean during the 1993-98 period [J]. Geophysical Research Letters, 2000, 27(11): 1659-1662. doi: 10.1029/1999GL011261
[64] Barkley H C, Cohen A L. Skeletal records of community-level bleaching in Porites corals from Palau [J]. Coral Reefs, 2016, 35(4): 1407-1417. doi: 10.1007/s00338-016-1483-3
[65] Elliot M, Welsh K, Chilcott C, et al. Profiles of trace elements and stable isotopes derived from giant long-lived Tridacna gigas bivalves: Potential applications in paleoclimate studies [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2009, 280(1-2): 132-142. doi: 10.1016/j.palaeo.2009.06.007
[66] Romanek C S, Grossman E L. Stable Isotope Profiles of Tridacna maxima as Environmental Indicators [J]. Palaios, 1989, 4(5): 402-413. doi: 10.2307/3514585
[67] Morimoto M, Abe O, Kayanne H, et al. Salinity records for the 1997-98 El Niño from Western Pacific corals [J]. Geophysical Research Letters, 2002, 29(11): 35-1-35-4.
[68] Warter V, Müller W. Daily growth and tidal rhythms in Miocene and modern giant clams revealed via ultra-high resolution LA-ICPMS analysis - A novel methodological approach towards improved sclerochemistry [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 465: 362-375. doi: 10.1016/j.palaeo.2016.03.019
[69] Yan H, Wang Y H, Sun L G. High resolution oxygen isotope and grayscale records of a medieval fossil giant clam (Tridacna gigas) in the South China Sea: physiological and paleoclimatic implications [J]. Acta Oceanologica Sinica, 2014, 33(8): 18-25. doi: 10.1007/s13131-014-0399-4
[70] Romanek C S, Jones D S, Williams D F, et al. Stable isotopic investigation of physiological and environmental changes recorded in shell carbonate from the giant clam Tridacna maxima [J]. Marine Biology, 1987, 94(3): 385-393. doi: 10.1007/BF00428244
[71] Fairbanks R G, Evans M N, Rubenstone J L, et al. Evaluating climate indices and their geochemical proxies measured in corals [J]. Coral Reefs, 1997, 16(5): S93-S100. doi: 10.1007/s003380050245
[72] 洪阿实, 洪鹰, 王庆春, 等. 1994年夏季南海东北部海水氧同位素分布特征[J]. 热带海洋, 1997, 16(2):82-90 HONG Ashi, HONG Ying, WANG Qingchun, et al. Distributive characteristics of O isotope of the northeastern South China Sea in the summer of 1994 [J]. Tropic Oceanology, 1997, 16(2): 82-90.
[73] Diaz H F, Hoerling M P, Eischeid J K. ENSO variability, teleconnections and climate change [J]. International Journal of Climatology, 2001, 21(15): 1845-1862. doi: 10.1002/joc.631
[74] Bellenger H, Guilyardi E, Leloup J, et al. ENSO representation in climate models: from CMIP3 to CMIP5 [J]. Climate Dynamics, 2014, 42(7-8): 1999-2018. doi: 10.1007/s00382-013-1783-z
[75] Versteegh E A A, Vonhof H B, Troelstra S R, et al. Seasonally resolved growth of freshwater bivalves determined by oxygen and carbon isotope shell chemistry [J]. Geochemistry, Geophysics, Geosystems, 2010, 11(8): Q08022.
[76] Woodroffe C D, Beech M R, Gagan M K. Mid-late Holocene El Niño variability in the equatorial Pacific from coral microatolls [J]. Geophysical Research Letters, 2003, 30(7): 1358.

 下载:
下载:








计量
- 文章访问数: 3476
- HTML全文浏览量: 823
- PDF下载量: 135


