
Changes of the upper water column at the 45°N North Atlantic since marine isotope stage 3
-
摘要: 北大西洋45°N区是北大西洋冰筏碎屑(IRD)带的中心区,其海洋沉积物包含高分辨率沉积环境和气候变化信息,对45°N区沉积记录的研究有利于反演末次冰期以来古海洋环境的变化。通过对岩心Hu71-377中IRD含量的统计、浮游有孔虫组合及氧和碳同位素(δ18O和δ13C)分析,重建了北大西洋45°N上层水体水团性质演化历史。结合AMS14C数据和氧同位素地层学,在氧同位素3期(MIS3)和2期(MIS2)中识别出5个Heinrich层,其中Heinrich 1、2和4层具有明显IRD峰值、Neogloboquadrina pachyderma高丰度和轻δ18O值特征,而Heinrich 3和5层的δ18O值未明显变轻。Heinrich 3和5层与Heinrich 1、2和4层的δ18O 差异可能反映了上层水体受融水输入的影响不同。δ13CN.incompta和δ13CN.pachyderma差值也反映了Heinrich事件期间混合层和温跃层的变化,它们的δ13C差值在Heinrich 1和2期间接近零,归因于强风驱动的海水垂向混合。而δ13CN.incompta和δ13CN.pachyderma差值在Heinrich 4和5期间增大,反映了季节性温跃层变浅,推测与北大西洋暖流增强有关。浮游有孔虫组合进一步反映了海洋上层水团性质, 特别是N. pachyderma和Neogloboquadrina incompta的相对丰度反映了MIS3期以来海表温度(SST)变化。
-
关键词:
- Heinrich事件 /
- 浮游有孔虫组合 /
- 碳氧同位素 /
- 上层水团变化 /
- 北大西洋
Abstract: The 45°N of North Atlantic is located at the central zone of the ice-rafted detritus (IRD) belt of the North Atlantic, where the marine sediments contain rich environmental and climatic information of high-resolution. The sedimentary records there are used for reconstruction of the pale-oceanic environment since the last glacial in this study. IRD contents, planktonic foraminiferal assemblages and their oxygen and carbon isotopes (δ18O and δ13C) from the core Hu71-377, are used as major tools. Combined with AMS14C dating and oxygen isotope stratigraphy, five Heinrich layers are identified in the MIS3 and MIS2, in which the Heinrich layer 1, 2 and 4 have obvious IRD peaks, high relative abundance of Neogloboquadrina pachyderma and light δ18O values, but no obvious light δ18O are observed in the Heinrich layer 3 and 5. The difference in δ18O between the Heinrich layers 3 and 5 and the Heinrich layers 1, 2 and 4 may suggest the impacts of melt water on the upper water column. Further, the offsets between δ13CN.incompta and δ13CN.pachyderma may also reflect the changes in the mixed layer and thermocline during the Heinrich events. The δ13C offsets were close to zero during Heinrich 1 and Heinrich 2, attributing to the vertical mixing of seawater driven by strong winds. And the δ13C offsets became larger during Heinrich 4 and Heinrich 5, indicating that the seasonal thermocline became shallower, which supports the inference of the penetration of the North Atlantic Current. What’s more, the planktonic foraminiferal assemblages may reflect the properties of the water masses in the upper water column, especially the relative abundance of N. pachyderma and Neogloboquadrina incompta may indicate the sea surface temperature (SST) changes during MIS3. -
北大西洋中纬度海域(40°~55°N)是北大西洋亚极地环流圈和亚热带环流圈相互作用的地带,是低温低盐的亚极地水和高温高盐的北大西洋暖流这两种不同性质水团的汇集地[1]。夏季北大西洋暖流向北增强,上层水体被亚热带温暖和贫营养水团所覆盖,导致水体分层和混合层变薄[2]。研究表明,冰期极地冷水从现代位置扩张至中纬度海域,洋流体系向南和向东移动,亚极地环流圈增强[3]。该现象归因于北大西洋周围冰盖的反复崩塌,产生大量融水,最终削弱北大西洋深层水的形成[4-5]。但是,目前对这些变化在北大西洋冰筏碎屑(Ice-rafted detritus,IRD)带内的影响程度仍不太清楚,有必要进行更深入的研究。
冷暖气候突变的Dansgaard-Oeschger事件(D-O事件)引发了末次冰期以来千年尺度气候变化的研究热潮[6-7]。格陵兰冰芯的研究最先报道了D-O事件,该事件还出现在许多全球气候记录中,如湖泊、洞穴、海洋沉积物[8-9]。Heinrich事件和D-O事件并不是两个孤立的气候演变过程,Heinrich事件发生在D-O事件的最冷格陵兰冰阶(Greenland Stadial),期间释放大量融水和碎屑,这些碎屑大多来自更新世晚期的北美劳伦泰冰盖[8,10-11]。通过北大西洋的IRD异常增加可以识别出Heinrich层,证实了冰盖的不稳定性和深层环流的减弱,导致北大西洋周围的气候极为寒冷[12-13]。
了解过去海洋表层温度(sea-surface temperature,SST)的变化对理解现代和过去海洋表层环流起着至关重要的作用。浮游有孔虫作为水体环境信息的载体,其分布受SST、盐度、营养盐等影响[2,14-15]。人们一直对使用浮游有孔虫的分布、相对丰度和稳定同位素(δ13C和δ18O)等方法探究上层水团变化十分感兴趣。CLIMAP(Climate Long-Range Investigation, Mapping and Prediction)首先利用浮游有孔虫转换函数评估了北大西洋末次冰盛期(Last Glacial Maximum,LGM)的SST[16-17]。Ruddiman和McIntyre[18-19]结合δ18O和有孔虫组合,重建了末次冰期上层水团性质特征和表层环流变化。Pflaumann等[20]改进了浮游有孔虫组合数据库,使有孔虫组合与SST具有更好的相关性,后被用于LGM表层水团和环流的评估[21]。Rashid和Boyle[22]研究IRD带南缘代表不同栖息深度的3种浮游有孔虫δ18O,评估了Heinrich事件期间上层水体的混合程度。然而,他们并未将有孔虫组合与δ18O结合在一起,对上层水团变化的了解并不充分[22-23]。此外,尚不清楚两种主要浮游有孔虫Neogloboquadrina incompta(N. incompta)和Neogloboquadrina pachyderma(N. pachyderma)的δ13C和δ18O之间的关系受上层水团变化的影响程度[24-25]。
本研究以Hu71022航次在北大西洋中脊45°N区采集的岩心Hu71-377为研究对象,旨在通过IRD、浮游有孔虫组合、δ18O和δ13C来探讨氧同位素3期(marineisotope stage,MIS3)以来北大西洋45°N区的上层水文环境变化。本研究还结合大洋钻探计划DSDP609[8]站位(Deep Sea Drilling Project)和格陵兰冰芯NGRIP[7](North Greenland Ice Core Project)的古海洋学记录,建立了Hu71-377年龄模型,为重建45°N区古海洋环境提供重要依据。
1. 材料与方法
1.1 材料
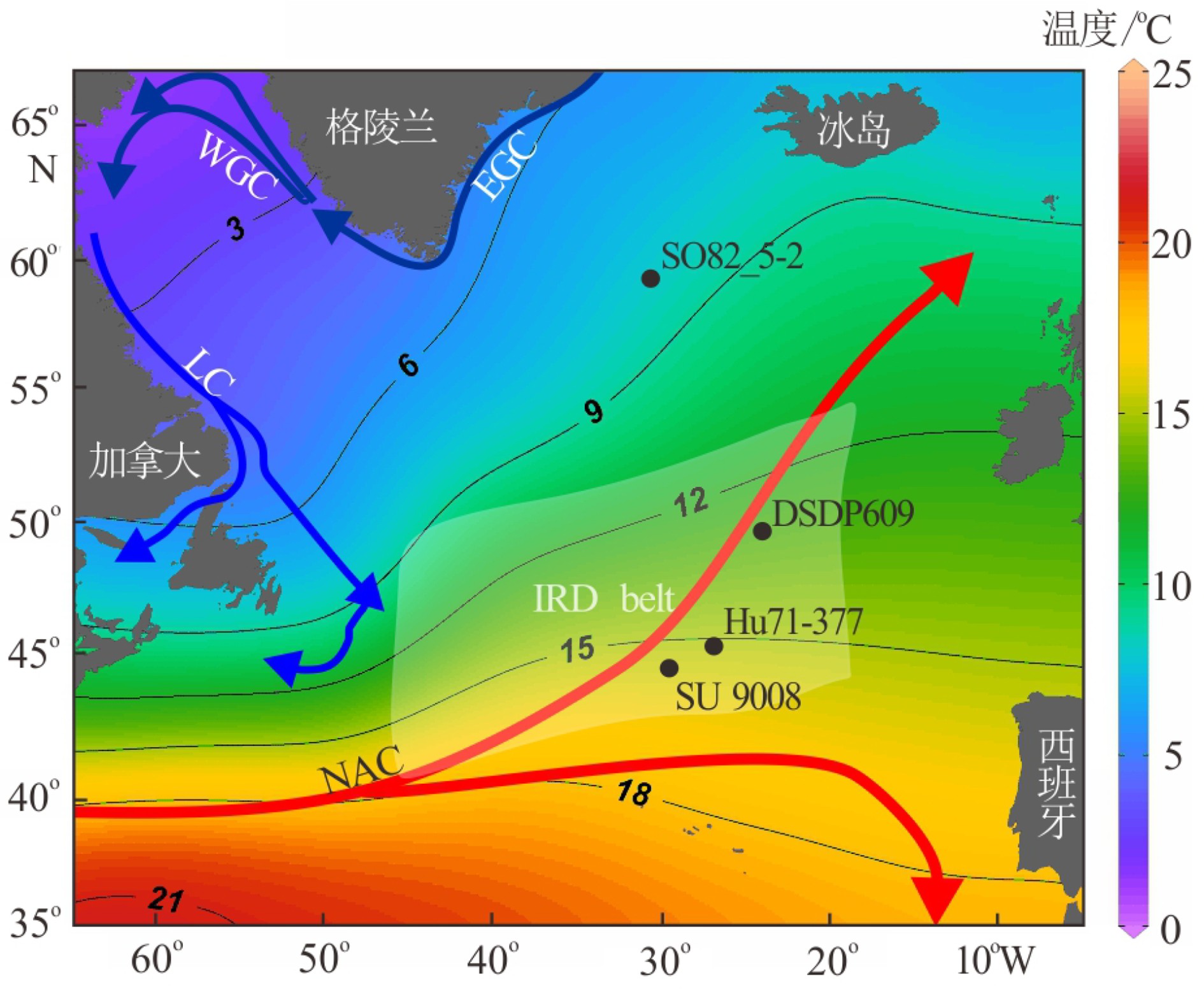
本次研究样品来自Hu71022航次获取的岩心Hu71-377(45.3181ºN、27.2561ºW,长11.5 m,水深2631 m)。该岩心位于IRD带内(图1)[26],保存完好且不存在沉积间断。Scott等[27]曾对该岩心进行间隔10 cm取样的分辨率研究,获得117个样品。后来,又在几个深度间隔补采了30个新样品以提高分辨率,共计147个样品。研究主要集中在岩心0 ~ 650 cm深度区间,仅使用83个样品进行分析。
![]() NAC—北大西洋暖流,LC—拉布拉多寒流,EGC—东格陵兰流,WGC—西格陵兰流,白色阴影表示IRD带。数据来源于World Ocean Atlas 18,由Ocean Data View绘制。Figure 1. Location map for Core Hu71-377 and related cores[8,28-29]and the distribution of currents and SST in the North AtlanticNAC—North Atlantic Current, LC—Labrador Current,EGC—East Greenland Current, WGC—West Greenland Current, white shade indicates the IRD belt. Data were download from World Ocean Atlas 18 and plotted by Ocean Data View.
NAC—北大西洋暖流,LC—拉布拉多寒流,EGC—东格陵兰流,WGC—西格陵兰流,白色阴影表示IRD带。数据来源于World Ocean Atlas 18,由Ocean Data View绘制。Figure 1. Location map for Core Hu71-377 and related cores[8,28-29]and the distribution of currents and SST in the North AtlanticNAC—North Atlantic Current, LC—Labrador Current,EGC—East Greenland Current, WGC—West Greenland Current, white shade indicates the IRD belt. Data were download from World Ocean Atlas 18 and plotted by Ocean Data View.1.2 方法
采集的样品首先在室温下浸泡24 h,直至样品在水中完全分散,再采用63 μm的筛子进行湿筛。用滤纸收集筛网中大于63 μm粒径的样品,放入60 ℃的烘箱中干燥24 h备用。
IRD鉴别及其含量:IRD是由浮冰携带的粗碎屑颗粒,当浮冰融化时,粗碎屑颗粒被释放到海水中并沿水柱沉降至海底;IRD含量可用来提供冰山数量、分布、冰川侵蚀和水温等信息。本研究IRD鉴别采用最常见和最直接的方法:对>150 µm粗碎屑颗粒(表面有尖锐棱角)进行计数[30]。
浮游有孔虫鉴定与相对丰度统计:筛出>150 μm粒径样品,并在分样器上进行缩分。7 ~ 8次缩分后将样品均匀洒在9×9网格的黑底托盘上,放在双目显微镜下观察并统计完整壳壁的有孔虫总数,当确定有孔虫总数超过300且小于600个时,进行浮游有孔虫鉴定和统计;若有孔虫总数不足300个或超过600个,则减少或增加1次缩分。此计数手段在海洋沉积学和古海洋学研究中是一种常用方法[8,22,31],本研究中浮游有孔虫鉴定参考Tolderlund和Bé [14]以及Schiebel和Hemleben [15]的方法。
δ18O、δ13C测试和壳体质量:从150 ~ 250 µm样品中分别挑选出20个表面无杂质的N. pachyderma和N. incompta完整壳体。用Metler Toledo微量天平称取这20个壳体总质量,最后计算出单个壳体质量。δ18O和δ13C测试在美国南佛罗里达大学地质实验室进行,使用配备Kiel Ⅲ碳酸盐前处理装置的Finnigan-MAT 252同位素质谱仪来测定。δ18O和δ13C测试的标准偏差分别是0.07‰和0.04‰[32]。
AMS14C测年:进行AMS14C测试的有孔虫包括N. pachyderma、Globigerina bulloides(G. bulloides)和Globorotalia inflata(G. inflata)。分别在岩心0 ~ 2 cm、68.5 ~ 69.5 cm、115 ~ 116 cm、256 ~ 257 cm和389 ~ 390 cm的样品中挑出壳体粒径为 150 ~ 250 µm的完整有孔虫壳体。每个样品需400 ~ 500个有孔虫壳体进行AMS14C测年,重6 ~ 8 mg。AMS14C测试在美国加利福尼亚大学欧文分校的W M Keck AMS实验室完成。
2. 结果
2.1 年龄模型
将表1中5个14C数据在MATLAB中运行Undatable[33],并采用Marine20校正曲线[34],将年龄模型控制在1.3至40.9 ka,即岩心0至390 cm。Undatable中执行“100000”仿真,得到的年龄-深度关系模型如图2a所示,取平均值计算沉积速率(图2b)。沉积速率在28.9 ~ 40.9 ka最高(13.2 cm/ka),随后逐渐降低并在1.3 ~ 12.9 ka降至最低(5.9 cm/ka)(图2b)。
表 1 岩心Hu71-377 14C年龄及日历年龄Table 1. 14C Age and Calendar Age of the core Hu71-377深度/cm 实验ID 14C年龄/a 测试材料 日历年龄/ka 0 ~ 2 UCI-212015 1930±15 G. inflata 1.333 68.5 ~ 69.5 UCI-214037 11545±25 G. inflata 12.860 115 ~ 116 UCI-23843 15840±60 N. pachyderma 18.174 256 ~ 257 UCI-212021 25610±80 G. inflata 28.887 389 ~ 390 UCI-214039 37120±230 G. bulloides 40.920 ![]() 图 2 岩心Hu71-377深度-年龄模型(a)和沉积速率(b)Figure 2. The depth-age model(a)and sedimentation rates of core Hu71-377(b)
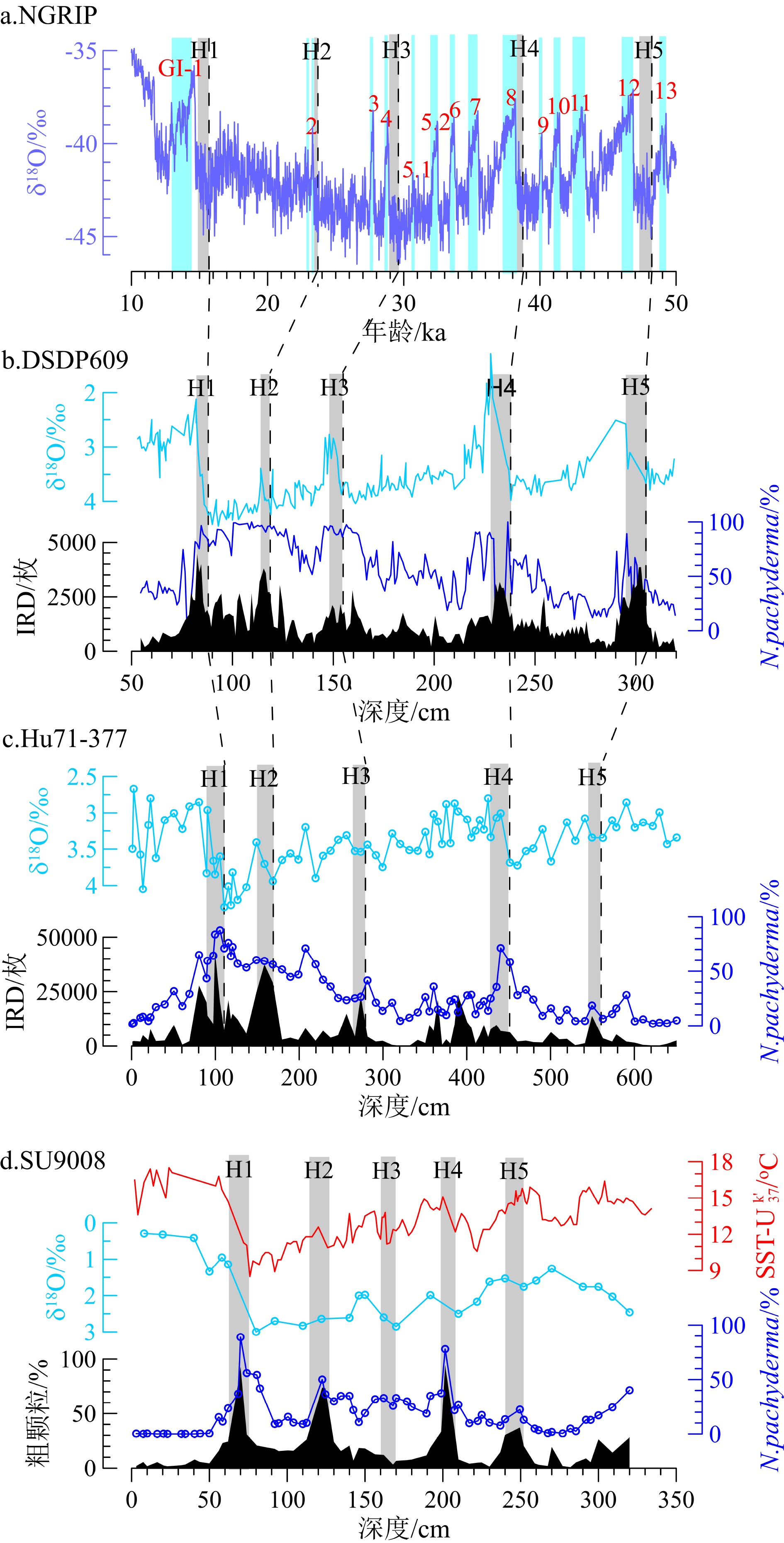
图 2 岩心Hu71-377深度-年龄模型(a)和沉积速率(b)Figure 2. The depth-age model(a)and sedimentation rates of core Hu71-377(b)通过与格陵兰冰芯NGRIP年代框架[7,35]和北大西洋深海沉积物中Heinrich层[36-37]对比,获得40 ka以上的年龄且校正Hu71-377近50 ka以来的年龄模型[38-39]。综合IRD、N. pachyderma丰度和δ18O值识别Hu71-377主要的Heinrich层(表2,图3),Heinrich层有以下特征:(1)有丰富的碎屑碳酸盐(IRD含量较高);(2)N. pachyderma丰度增加和δ18O变轻[8,22]。据此标志特征,岩心Hu71-377和DSDP609中Heinrich 1、2和4沉积层具有相似的IRD、N. pachyderma丰度和δ18O变化特征。
表 2 NGRIP冰芯、DSDP609岩心和Hu71-377岩心的Heinrich事件相关控制点Table 2. Heinrich events in the ice core NGRIP, core DSDP609 and core Hu71-377![]()
综上,Hu71-377的年龄模型如图4所示,时间序列大致分为:0~90 cm为MIS1(14.3 ka至现代,包括全新世10 ka至今[40],对应0 ~ 50 cm);91 本文针对滨海地区地下咸水的来源和演化问题,以江苏盐城为研究区,通过水化学,同位素的方法,对研究区的地下水做了大量现场调查和分析研究工作,具有一定的理论意义和应用价值。本次修改在之前的提出的问题上做了较多改正,文章质量具有提高,可刊用。 256 cm为MIS2,对应时间序列14.3 ~ 28.9 ka(LGM发生在19 ~ 26.5 ka[41],对应126 ~ 219 cm);MIS3期(28.9 ~ 51.3 ka)位于岩心257 ~ 650 cm。参考Griem等[38]对MIS3期的划分,本研究中MIS3期可划分为暖期(本研究中为40 ~ 51.3 ka)和冷期(28.9 ~ 40 ka)。
![]() “?”表示岩心SO82_5-2中Heinrich 2的年龄模型存在不确定性,因为van Kreveld等[29]未对Heinrich 2有详细定义,并且与其他Heinrich层相比,Heinrich 2中δ18O和δ13C特征不明显。Figure 4. IRD, relative abundance of foraminifera, δ18O and δ13C, and weights of foraminifera in core Hu71-377 and their correlation with δ18O and δ13C from core SO82_5-2[29] and core DSDP609[37]“?”represents an uncertain age model of Heinrich 2 in core SO82_5-2, since van Kreveld et al.(2000) [29] did not provide detailed definition for Heinrich 2 and the δ18O and δ13C in Heinrich 2 in SO82_5-2 are not well correlated with other Heinrich layers
“?”表示岩心SO82_5-2中Heinrich 2的年龄模型存在不确定性,因为van Kreveld等[29]未对Heinrich 2有详细定义,并且与其他Heinrich层相比,Heinrich 2中δ18O和δ13C特征不明显。Figure 4. IRD, relative abundance of foraminifera, δ18O and δ13C, and weights of foraminifera in core Hu71-377 and their correlation with δ18O and δ13C from core SO82_5-2[29] and core DSDP609[37]“?”represents an uncertain age model of Heinrich 2 in core SO82_5-2, since van Kreveld et al.(2000) [29] did not provide detailed definition for Heinrich 2 and the δ18O and δ13C in Heinrich 2 in SO82_5-2 are not well correlated with other Heinrich layers2.2 IRD和浮游有孔虫组合变化
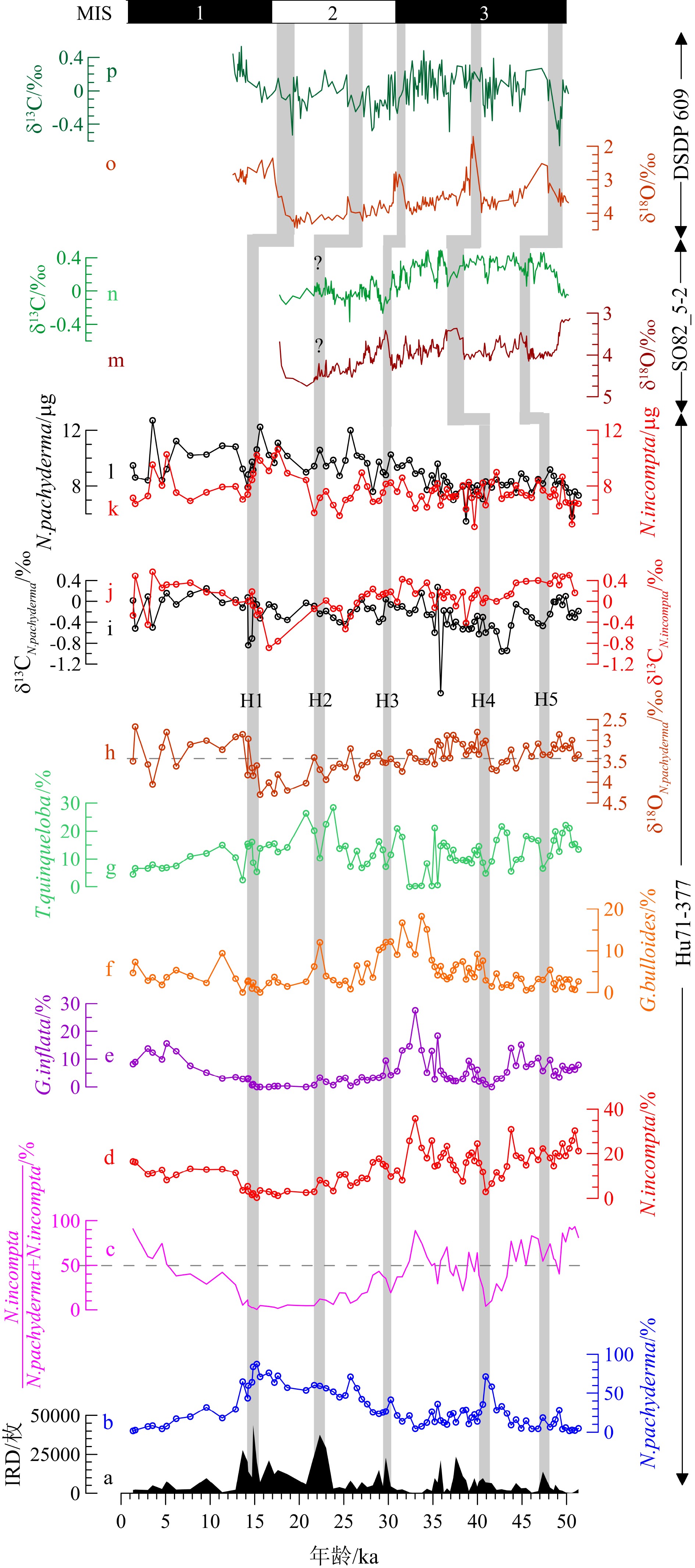
Hu71-377的IRD含量变化很大,范围为128~43552枚(图4a)。MIS3期IRD含量最低,IRD峰值(>10000枚)出现在Heinrich 5、Heinrich 4、37.6 ka、35.9 ka 和Heinrich 3。MIS2期IRD含量最高,IRD最大值出现在Heinrich 2。14.8 ka IRD峰值对应Heinrich 1。MIS1以来IRD含量减少,无明显峰值。
浮游有孔虫主要由N. pachyderma、N. incompta、G. bulloides、G. inflata、Turborotalita quinqueloba(T. quinqueloba)组成,其中N. pachyderma最丰富(图4b、4d-g)。全新世还有少量粉色和白色Globigerinoides ruber(G. ruber)。MIS3期N. incompta、G. bulloides和G. inflata丰度最高,在31~35 ka出现最大值35.84%、18.24%和27.53%;而N. pachyderma相对丰度较低,仅在Heinrich 4出现极大值。N. pachyderma在MIS2期逐渐增加,而N. incompta、G. bulloides和G. inflata相对丰度偏低。MIS1以来N. pachyderma和T. quinqueloba减少,N. incompta、G. bulloides和G. inflata相对丰度增加。
2.3 N. pachyderma和N. incompta的δ18O和δ13C
Hu71-377中N. pachyderma的δ18O为2.67‰ ~ 4.30‰,平均3.39‰(图4h)。MIS3期δ18O偏轻,在Heinrich 4明显变轻(2.80‰)。Heinrich 4后δ18O逐渐变重,至Heinrich 1前δ18O达到最大值(4.30‰),期间Heinrich 2有极小值(3.41‰)。Heinrich 1期间δ18O显著变轻。在MIS1期δ18O偏轻,平均3.23‰。N. pachyderma和N. incompta的δ13C(分别用δ13CN.pachyderma和δ13CN.incompta表示)分别在–1.76 ~ 0.27‰和–0.89 ~ 0.57‰变化(图4i、4j)。MIS3期δ13CN.incompta比δ13CN.pachyderma重,平均为0.18‰和−0.32‰。除17 ka有2个δ13CN.incompta异常低值外,MIS2和MIS1期δ13CN.incompta与δ13CN.pachyderma相似。所有Heinrich事件中,Heinrich 4和5期间显示较大的δ13C差值,为0.65‰和0.80‰,而Heinrich 1和2期间δ13C差值接近零。
2.4 N. pachyderma和N. incompta壳体质量
Hu71-377中N. pachyderma和N. incompta壳体质量分别为5.47 ~ 12.70 µg/个和5.06 ~ 10.65 µg/个,平均值为8.89 µg/个和7.65 µg/个(图4k、4l)。35 ~ 51.3 ka和Heinrich 1期间N. incompta与N. pachyderma壳体质量相同,其中N. pachyderma质量在Heinrich 1期间增加34.6%。LGM和全新世N. incompta壳体质量偏轻,N. pachyderma和N. incompta间的质量差为3 µg/个(图4k、4l)。
3. 讨论
3.1 Heinrich事件
与北大西洋岩心DSDP609和SU9008数据对比,Hu71-377中IRD分布模式与北大西洋中纬度(40°~55°N)岩心[10,28,36]记录相符(图3)。岩心Hu71-377中识别了5个位于MIS3至MIS2期的Heinrich层,其中Heinrich 1、2和4 层都具有与IRD带内Heinrich层相一致的高IRD含量、高N. pachyderma丰度及轻δ18O特征。值得注意的是,Hu71-377中Heinrich 4期间δ18O减少了1.0‰,与北大西洋43°~50°N岩心中δ18O减少0.5‰ ~ 1.7‰相符[42];但是Hu71-377中Heinrich 4层没有与其他岩心一样出现异常的IRD值,尚不清楚其具体原因。
研究表明,北大西洋东部岩心Heinrich 3层中缺少碎屑碳酸盐记录(IRD含量较低)[43],而其他Heinrich层中都有明显的IRD峰值特征,这与Hudson Strait释放的冰山有关[10-11]。但是,在Hu71-377和DSDP609的IRD记录中Heinrich 3层都有一个狭窄的IRD高峰,反映了Heinrich 3期间研究区发生短时间的冰盖扩张。此外,与岩心SU9008[28]记录相似,岩心Hu71-377中Heinrich 3层也没有较高的N. pachyderma丰度特征,可能反映了Heinrich 3期间极地锋在短时间内南移至45°N。
Heinrich事件期间融水释放使得δ18O变轻[22,44],但Hu71-377中Heinrich 3和5层未观察到与Heinrich 1、2和4层一样的δ18O变化特征,一方面可能是低分辨率采样所致(间隔5 ~ 10 cm);另一方面,与DSDP609相比,位于IRD带西南位置的Hu71-377易受到亚热带环流圈和亚极地环流圈的交替影响。因为冰期亚热带环流圈和亚极地环流圈的南缘以及最大冰山融化带[26]是变动的,它们在Heinrich期间没有固定位置[31]。在Heinrich 1、2和4期间,Hu71-377站位受冰山融水的影响更强烈,而Heinrich 3和5期间可能有更多的暖水流入,从而减弱了冰山融水的影响,这是δ18O 在Heinrich 3和5期间与Heinrich 1、2和4期间变化不同的原因之一。在附近岩心SU9008中发现了相似的δ18O 差异,Missiaen等使用浮游有孔虫组合报道了近45 ka以来SU9008站位的SST变化[45],发现Heinrich 2和4开始时SST下降,随后SST逐渐升高,表明北大西洋暖水在Heinrich 2和4后输入SU9008站位。而Heinrich 3期间没有明显的SST变化,这意味着 Heinrich 3期间北大西洋暖水盛行。因此,SU9008站位Heinrich 3、5期间不同于Heinrich 2、4期间SST的变化可以归因于Heinrich 3、5期间北大西洋暖流输入,海表热量增加,该现象可能同样发生在Hu71-377站位。此外,通过比较岩心CH69-K09和CHN82-20中3种浮游有孔虫G. bulloides(0 ~ 30 m)、N. incompta(30 ~ 100 m)和N. pachyderma(100 ~ 250 m)的δ18O值,Rashid和Boyle认为Heinrich事件期间上层水体混合深度不同,从而导致δ18O的变化不同[22]。本研究中仅记录了温跃层栖息的N. pachyderma(100 ~ 250 m)在Heinrich 3和5期间无明显δ18O轻值,因此不能排除海水混合影响了最上层水体(温跃层以上)的可能性。
3.2 有孔虫组合及其古海洋学意义
在极地至亚极地海区,N. pachyderma常见于温跃层,其较高的通量和相对丰度与低SST相对应[14,46]。Govin等将40°N以北的N. pachyderma相对丰度与SST相关联:N. pachyderma相对丰度为80%相当于~4 ℃[46],且N. pachyderma丰度随SST增加而减少,这与Pflaumann等的转换函数结果一致[20]。N. incompta是常见于过渡带至亚极地的混合层种,并且在研究区其相对丰度变化是北大西洋暖流流入的指标之一[47-48]。由此可知,N. pachyderma和N. incompta均受SST影响,表现出截然不同的生态意义:N. pachyderma代表较冷的水文环境(<9 ℃)[47];而N. incompta代表相对温暖的水文环境(10 ~ 18 ℃)[14,49]。研究50°N ~ 65°S海域浮游有孔虫对SST的敏感性时,Zaric等[47]提出N. incompta在约9 ℃超过N. pachyderma的相对丰度,这与Govin等[46]的研究相符。岩心Hu71-377的数据显示,N. pachyderma和N. incompta占优势,它们的相对丰度呈镜像关系,N. incompta与N. pachyderma丰度的比值[50]可能反映与水团变迁有关的SST变化(图4c),该结果与附近岩心SU9008[51](图3d)和T88-9p[52]中利用Uk’ 37温度指标得到的SST变化趋势一致。当它们的比值高于50%时,SST高于 9 ℃,反之则相反。MIS2期N. incompta与N. pachyderma丰度的比值低于50%,反映了极地水团侵入导致SST降低(<9 ℃),这主要与极地锋南移[53]有关。此外,T. quinqueloba相对丰度随N. pachyderma同步增加,SU9003站位记录了相同的浮游有孔虫组合变化[31],反映了极地锋在MIS2期至少南移至40°N区,因此北大西洋40°N以北上层水体被极地冷水覆盖,有利于N. pachyderma和T. quinqueloba生长。相反,它们的比值在MIS3期暖期高于50%,还在全新世逐渐增加至85%,都暗示了相对温暖的海洋环境,反映北大西洋暖流流入,导致SST较高。
值得注意的是,N. incompta丰度最大值出现在31 ~ 35 ka, G. bulloides和G. inflata丰度也相应增大(图4d-f)。由于北大西洋45°N区N. incompta、G. bulloides和G. inflata的相对丰度大小反映与北大西洋暖流流入[48]有关的上层水体变化,因此该期间上层水体变化可能具有强烈的季节性特征。Jonkers等[30]报道了北大西洋暖流流入和有孔虫季节性生长,可能导致Heinrich 4后SST升高。由此,我们推测研究区北大西洋暖流在31 ~ 35 ka并未减弱,且在夏季增强,导致这些浮游有孔虫在温暖季节勃发,因此其相对丰度增加。
3.3 浮游有孔虫碳氧同位素记录与上层水体性质
岩心Hu71-377在全新世、MIS3期暖期的D-O事件以及Heinrich期间的δ18O值轻于其平均值,反映上层水体扰动,相似的δ18O特征也在北大西洋沉积记录中发现。浮游有孔虫δ18O主要受SST和盐度影响,因此SST升高和冰融水导致的盐度降低,都将导致N. pachyderma的δ18O变轻[22,44]。结合岩心Hu71-377的浮游有孔虫组合显示,N. pachyderma和N. incompta的相对丰度比值在全新世和MIS3期暖期(主要为D-O事件)较高,反映SST较高,这与较轻的δ18O值一致。甚至有孔虫壳体δ18O与SST的相关性表明,当SST升高1 ℃时,N. pachyderma的δ18O降低0.25‰[54]。盐度也是影响δ18O的重要因素,在研究区可以排除河流淡水注入的可能性,但是D-O事件和Heinrich事件期间不同来源的淡水注入也会使得δ18O变轻[4,42]。特别是Heinrich事件大量冰融水注入研究区,导致海水盐度显著降低,同时N. pachyderma栖息在较深的水体中[14],受盐跃层的影响,其δ18O变轻。但是,Hu71-377站位的δ18O、IRD和N. pachyderma相对丰度的多变(图3),说明了并非所有的Heinrich事件期间冰融水同等程度的影响上层水体结构。
底栖有孔虫δ13C常被用来示踪深水团演化和重建过去海洋环流[55-56],如底栖有孔虫δ13C在LGM变轻,反映北大西洋深层水输入减弱。浮游有孔虫δ13C和底栖有孔虫δ13C反映了不同性质的水团和海洋环境。浮游有孔虫δ13C是海水中总溶解无机碳(∑CO2)δ13C的函数[57],海水∑CO2的δ13C主要受控于:(1)有机质形成和降解所引起的δ13C变化[57-58],如光合作用、营养盐等;(2)海气交换引起的δ13C变化,如风的扰动引起强烈的海气交换[59]。此外,大量冰融水注入也可以解释δ13C变轻[44,60],如北大西洋岩心(包括DSDP609和SO82_5-2,图4m-p)中N. pachyderma的δ13C在Heinrich期间明显变轻[29,37]。尽管δ13CN.pachyderma也显示δ13C轻值,但是在Hu71-377站位冰融水可能对δ13C的影响不大。综合δ13CN.pachyderma和δ13CN.incompta以及浮游有孔虫组合变化,发现Heinrich事件期间G. bulloides、G. inflata和T. quinqueloba等浮游有孔虫的相对丰度较低,而N. incompta和N. pachyderma仍占主导地位,因此N. incompta和N. pachyderma代表了这些浮游有孔虫生长的上层水体环境,它们的δ13C主要反映该期间海水∑CO2的δ13C变化,即δ13CN.pachyderma和δ13CN.incompta直接反映了Heinrich事件期间研究区的上层水体环境。
由于表层海水的光合作用优先吸收较轻的12C,导致表层海水∑CO2的δ13C变重。有机质在沉降过程中(生物泵效应)被降解,使海水∑CO2的δ13C随水深加大而变轻[55], 这样导致混合层种N. incompta的δ13C重于温跃层种N. pachyderma。因此,在本研究中δ13CN.incompta和δ13CN.pachyderma差值被用来反映Heinrich事件期间温跃层的深浅。Heinrich 1和2期间δ13CN.incompta和δ13CN.pachyderma差值减小,说明温跃层变深。除冰融水注入导致混合层变深外,该现象还可能是由于MIS2冷期强风驱动引起的海水垂向混合[22,59],使得δ13CN.incompta随混合层加深而变轻,所以δ13CN.incompta和δ13CN.pachyderma差值减小。但是,Heinrich 4和5期间δ13CN.incompta和δ13CN.pachyderma差值增大,表明季节性温跃层变浅,推测是由于夏季北大西洋暖流增强导致的。该结果与Keigwin 和Boyle的发现一致:当温跃层变浅或夏季水体分层时,N. incompta和N. pachyderma的δ13C差值较大[56]。
在西北太平洋有一项研究,李铁刚等[61]分析了西北太平洋过去40 ka以来混合层种G. ruber和温跃层种Neogloboquadrina dutertrei的δ13C,通过δ13C差值反映的营养盐梯度,探讨了与黑潮有关的对马暖流的演化和变动。如黑潮的存在和陆源淡水输入引起营养盐梯度的变化,使δ13C差值增大。混合层和温跃层的营养盐梯度是否可以用来反映贫营养的北大西洋暖流的强弱尚不清楚。倘若这一假设能够应用于北大西洋暖流的演化,那么Heinrich 4和5期间北大西洋暖流夏季增强时,引起研究区混合层和温跃层的营养盐梯度增大,此时δ13CN.incompta和δ13CN.pachyderma差值增大也可能是温暖而贫营养的水团在近表层聚集,δ13CN.incompta变重所致。
此外,研究表明同一种内有孔虫壳体质量和壳体大小的差异会引起δ13C不同程度的差异[56,62-63]。Hu71-377中N. pachyderma壳体质量在Heinrich 1期间增加34.6%,而δ13CN.pachyderma不变(图4i、4l),这与Kohfeld等研究结果相符[24]。N. incompta壳体质量与δ13CN.incompta在LGM呈正相关而在全新世呈负相关(图4j、4k),说明了壳体质量可能会引起壳体δ13C与海水∑CO2的δ13C偏移。因此,在讨论上述δ13C差值反映上层水体环境时,不排除Heinrich期间壳体质量增加可能导致δ13CN.incompta变重。然而,我们尚不知晓此类偏移程度具体与哪些因素有关,该结论还有待商榷。
4. 结论
(1)北大西洋中纬海域岩心Hu71-377识别了5个位于MIS3—MIS2的Heinrich层。其中Heinrich 1、2和4层都具有IRD峰值、N. pachyderma高丰度及δ18O轻值特征,这与IRD带内Heinrich层的特征一致,但是Heinrich 3和5层中δ18O值没有明显变轻,该期间可能有更多的暖水输入,从而减弱了冰山融水的影响。
(2)浮游有孔虫以N. pachyderma和N. incompta为主,其相对丰度的比值与SST相关,可能反映与极地锋扩张有关的冷水团输入和与北大西洋暖流增强有关的暖水团输入。N. incompta、G. bulloides和G. inflata相对丰度最大值出现在31 ~ 35 ka,该现象具有较强的季节性特征,可能反映夏季北大西洋暖流增强。
(3)浮游有孔虫组合和δ13C结果表明,Heinrich事件期间N. incompta和N. pachyderma占主导地位,通过δ13CN.incompta和δ13CN.pachyderma差值主要反演研究区上层水体环境。Heinrich 1和2期间差值减小,可能受温跃层变深或强风驱动的海水垂向混合控制,而Heinrich 4和5期间差值增大,说明季节性温跃层变浅,推测与北大西洋暖流增强和夏季水体分层有关。
致谢:本研究的样品是加拿大自然科学与工程研究委员会(Natural Sciences and Engineering Research Council of Canada,NSERC)资助采集,由David Scott(Scott教授隶属于加拿大哈利法斯特戴尔豪西大学)提供岩心Hu71-377样品,对此深表谢意。感谢国家自然科学基金(41776064)资助,感谢加拿大大西洋沿岸地区机署(Atlantic Canada Opportunities Agency, ACOA)以及加拿大纽芬兰和拉布拉多地方政府的支持。感谢中国科学院南海海洋研究所黎刚老师、上海海洋大学罗敏老师对文章进行指点和润色。
-
![]()
图 1 北大西洋Hu71-377与相关岩心[8,28-29]的位置以及洋流和SST分布
NAC—北大西洋暖流,LC—拉布拉多寒流,EGC—东格陵兰流,WGC—西格陵兰流,白色阴影表示IRD带。数据来源于World Ocean Atlas 18,由Ocean Data View绘制。
Figure 1. Location map for Core Hu71-377 and related cores[8,28-29]and the distribution of currents and SST in the North Atlantic
NAC—North Atlantic Current, LC—Labrador Current,EGC—East Greenland Current, WGC—West Greenland Current, white shade indicates the IRD belt. Data were download from World Ocean Atlas 18 and plotted by Ocean Data View.
![]()
图 2 岩心Hu71-377深度-年龄模型(a)和沉积速率(b)
Figure 2. The depth-age model(a)and sedimentation rates of core Hu71-377(b)
![]()
![]()
图 4 北大西洋Hu71-377岩心IRD、有孔虫相对丰度、δ18O和δ13C、壳体质量等指标与岩心SO82_5-2[29] 和DSDP609[37]的δ18O和δ13C指标
“?”表示岩心SO82_5-2中Heinrich 2的年龄模型存在不确定性,因为van Kreveld等[29]未对Heinrich 2有详细定义,并且与其他Heinrich层相比,Heinrich 2中δ18O和δ13C特征不明显。
Figure 4. IRD, relative abundance of foraminifera, δ18O and δ13C, and weights of foraminifera in core Hu71-377 and their correlation with δ18O and δ13C from core SO82_5-2[29] and core DSDP609[37]
“?”represents an uncertain age model of Heinrich 2 in core SO82_5-2, since van Kreveld et al.(2000) [29] did not provide detailed definition for Heinrich 2 and the δ18O and δ13C in Heinrich 2 in SO82_5-2 are not well correlated with other Heinrich layers
表 1 岩心Hu71-377 14C年龄及日历年龄
Table 1 14C Age and Calendar Age of the core Hu71-377
深度/cm 实验ID 14C年龄/a 测试材料 日历年龄/ka 0 ~ 2 UCI-212015 1930±15 G. inflata 1.333 68.5 ~ 69.5 UCI-214037 11545±25 G. inflata 12.860 115 ~ 116 UCI-23843 15840±60 N. pachyderma 18.174 256 ~ 257 UCI-212021 25610±80 G. inflata 28.887 389 ~ 390 UCI-214039 37120±230 G. bulloides 40.920  下载: 导出CSV
下载: 导出CSV
表 2 NGRIP冰芯、DSDP609岩心和Hu71-377岩心的Heinrich事件相关控制点
Table 2 Heinrich events in the ice core NGRIP, core DSDP609 and core Hu71-377
下载: 导出CSV
-
[1] Lozier M S, Li F, Bacon S, et al. A sea change in our view of overturning in the subpolar North Atlantic [J]. Science, 2019, 363(6426): 516-521. doi: 10.1126/science.aau6592
[2] Cléroux C, Cortijo E, Anand P, et al. Mg/Ca and Sr/Ca ratios in planktonic foraminifera: Proxies for upper water column temperature reconstruction [J]. Paleoceanography and Paleoclimatology, 2008, 23(3): PA3214.
[3] Holliday N P, Bersch M, Berx B, et al. Ocean circulation causes the largest freshening event for 120 years in eastern subpolar North Atlantic [J]. Nature Communications, 2020, 11: 585. doi: 10.1038/s41467-020-14474-y
[4] Bagniewski W, Meissner K J, Menviel L. Exploring the oxygen isotope fingerprint of Dansgaard-Oeschger variability and Heinrich events [J]. Quaternary Science Reviews, 2017, 159: 1-14. doi: 10.1016/j.quascirev.2017.01.007
[5] Zhang X, Prange M. Stability of the Atlantic overturning circulation under intermediate (MIS3) and full glacial (LGM) conditions and its relationship with Dansgaard-Oeschger climate variability [J]. Quaternary Science Reviews, 2020, 242: 106443. doi: 10.1016/j.quascirev.2020.106443
[6] Dansgaard W, Johnsen S J, Clausen H B, et al. Evidence for general instability of past climate from a 250-kyr ice-core record [J]. Nature, 1993, 364(6434): 218-220. doi: 10.1038/364218a0
[7] Rasmussen S O, Bigler M, Blockley S P, et al. A stratigraphic framework for abrupt climatic changes during the last glacial period based on three synchronized Greenland ice-core records: refining and extending the INTIMATE event stratigraphy [J]. Quaternary Science Reviews, 2014, 106: 14-28. doi: 10.1016/j.quascirev.2014.09.007
[8] Bond G C, Heinrich H, Broecker W S, et al. Evidence for massive discharges of icebergs into the North Atlantic Ocean during the last glacial period [J]. Nature, 1992, 360(6401): 245-249. doi: 10.1038/360245a0
[9] Voelker A H L. Global distribution of centennial-scale records for Marine Isotope Stage (MIS) 3: a database [J]. Quaternary Science Reviews, 2002, 21(10): 1185-1212. doi: 10.1016/S0277-3791(01)00139-1
[10] Heinrich H. Origin and consequences of cyclic ice rafting in the northeast Atlantic Ocean during the past 130, 000 years [J]. Quaternary Research, 1988, 29(2): 142-152. doi: 10.1016/0033-5894(88)90057-9
[11] Hemming S R. Heinrich events: Massive late Pleistocene detritus layers of the North Atlantic and their global climate imprint [J]. Reviews of Geophysics, 2004, 42(1): RG1005.
[12] Broecker W. Massive iceberg discharges as triggers for global climate change [J]. Nature, 1994, 372(6505): 421-424. doi: 10.1038/372421a0
[13] Guo C C, Nisancioglu K H, Bentsen M, et al. Equilibrium simulations of Marine Isotope Stage 3 climate [J]. Climate of the Past, 2019, 15(3): 1133-1151. doi: 10.5194/cp-15-1133-2019
[14] Tolderlund D S, Be A W H. Seasonal distribution of planktonic foraminifera in the western North Atlantic [J]. Micropaleontology, 1971, 17(3): 297-329. doi: 10.2307/1485143
[15] Schiebel R, Hemleben C. Classification and taxonomy of extant planktic foraminifers[C]//Planktic Foraminifers in the Modern Ocean. Berlin: Springer, 2017: 11-110.
[16] McIntyre A, Kipp N G, Bé A W H, et al. Glacial North Atlantic 18, 000 years ago: A CLIMAP reconstruction[M]//Cline R M, Hays D J. Investigation of Late Quaternary Paleoceanography and Paleoclimatology. Boulder, Colorado: Geological Society of America, 1976: 43-76.
[17] CLIMAP Project Members. The surface of the ice-age earth [J]. Science, 1976, 191(4232): 1131-1137. doi: 10.1126/science.191.4232.1131
[18] Ruddiman W F, McIntyre A. The mode and mechanism of the last deglaciation: Oceanic evidence [J]. Quaternary Research, 1981, 16(2): 125-134. doi: 10.1016/0033-5894(81)90040-5
[19] Ruddiman W F, Raymo M E, Martinson D G, et al. Pleistocene evolution: northern hemisphere ice sheets and North Atlantic Ocean [J]. Paleoceanography and Paleoclimatology, 1989, 4(4): 353-412.
[20] Pflaumann U, Duprat J, Pujol C, et al. SIMMAX: A modern analog technique to deduce Atlantic sea surface temperatures from planktonic foraminifera in deep-sea sediments [J]. Paleoceanography and Paleoclimatology, 1996, 11(1): 15-35.
[21] Sarnthein M, Pflaumann U, Weinelt M. Past extent of sea ice in the northern North Atlantic inferred from foraminiferal paleotemperature estimates [J]. Paleoceanography and Paleoclimatology, 2003, 18(2): 1047.
[22] Rashid H, Boyle E A. Mixed-layer deepening during Heinrich events: a multi-planktonic foraminiferal δ18O approach [J]. Science, 2007, 318(5849): 439-441. doi: 10.1126/science.1146138
[23] Rashid H, Boyle E A. Response to comment on “Mixed-layer deepening during Heinrich events: a multi-planktonic foraminiferal δ18O approach” [J]. Science, 2008, 320(5880): 1161.
[24] Kohfeld K E, Fairbanks R G, Smith S L, et al. Neogloboquadrina pachyderma (sinistral coiling) as paleoceanographic tracers in polar oceans: evidence from northeast water polynya plankton tows, sediment traps, and surface sediments [J]. Paleoceanography and Paleoclimatology, 1996, 11(6): 679-699.
[25] Brummer G J A, Metcalfe B, Feldmeijer W, et al. Modal shift in North Atlantic seasonality during the last deglaciation [J]. Climate of the Past, 2020, 16(1): 265-282. doi: 10.5194/cp-16-265-2020
[26] Ruddiman W F. Late Quaternary deposition of ice-rafted sand in the subpolar North Atlantic (lat 40° to 65°N) [J]. GSA Bulletin, 1977, 88(12): 1813-1827. doi: 10.1130/0016-7606(1977)88<1813:LQDOIS>2.0.CO;2
[27] Scott D B, Baki V, Younger C D, et al. Empirical method for measuring seasonality in deep-sea cores [J]. Geology, 1986, 14(8): 643-646. doi: 10.1130/0091-7613(1986)14<643:EMFMSI>2.0.CO;2
[28] Grousset F E, Labeyrie L, Sinko J A, et al. Patterns of ice-rafted detritus in the glacial north Atlantic (40-55°N) [J]. Paleoceanography and Paleoclimatology, 1993, 8(2): 175-192.
[29] Van Kreveld S, Sarnthein M, Erlenkeuser H, et al. Potential links between surging ice sheets, circulation changes, and the Dansgaard-Oeschger cycles in the Irminger Sea, 60-18 kyr [J]. Paleoceanography and Paleoclimatology, 2000, 15(4): 425-442.
[30] Jonkers L, Moros M, Prins M A, et al. A reconstruction of sea surface warming in the northern North Atlantic during MIS 3 ice-rafting events [J]. Quaternary Science Reviews, 2010, 29(15-16): 1791-1800. doi: 10.1016/j.quascirev.2010.03.014
[31] Chapman M R, Shackleton N J, Duplessy J C. Sea surface temperature variability during the last glacial-interglacial cycle: assessing the magnitude and pattern of climate change in the North Atlantic [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 157(1-2): 1-25. doi: 10.1016/S0031-0182(99)00168-6
[32] Rashid H, Piper D J W, Drapeau J, et al. Sedimentology and history of sediment sources to the NW Labrador Sea during the past glacial cycle [J]. Quaternary Science Reviews, 2019, 221: 105880. doi: 10.1016/j.quascirev.2019.105880
[33] Lougheed B C, Obrochta S P. A rapid, deterministic age-depth modeling routine for geological sequences with inherent depth uncertainty [J]. Paleoceanography and Paleoclimatology, 2009, 34(1): 122-133.
[34] Heaton T J, Köhler P, Butzin M, et al. Marine20-the marine radiocarbon age calibration curve (0-55,000 cal BP) [J]. Radiocarbon, 2020, 62(4): 779-820. doi: 10.1017/RDC.2020.68
[35] Seierstad I K, Abbott P M, Bigler M, et al. Consistently dated records from the Greenland GRIP, GISP2 and NGRIP ice cores for the past 104 ka reveal regional millennial-scale δ18O gradients with possible Heinrich event imprint [J]. Quaternary Science Reviews, 2014, 106: 29-46. doi: 10.1016/j.quascirev.2014.10.032
[36] Bond G, Broecker W, Johnsen S, et al. Correlations between climate records from North Atlantic sediments and Greenland ice [J]. Nature, 1993, 365(6442): 143-147. doi: 10.1038/365143a0
[37] Obrochta S P, Miyahara H, Yokoyama Y, et al. A re-examination of evidence for the North Atlantic “1500-year cycle” at site 609 [J]. Quaternary Science Reviews, 2012, 55: 23-33. doi: 10.1016/j.quascirev.2012.08.008
[38] Griem L, Voelker A H L, Berben S M P, et al. Insolation and glacial meltwater influence on sea-ice and circulation variability in the northeastern Labrador Sea during the last glacial period [J]. Paleoceanography and Paleoclimatology, 2019, 34(11): 1689-1709. doi: 10.1029/2019PA003605
[39] Lisiecki L E, Stern J V. Regional and global benthic δ18O stacks for the last glacial cycle [J]. Paleoceanography and Paleoclimatology, 2016, 31(10): 1368-1394.
[40] Came R E, Oppo D W, McManus J F. Amplitude and timing of temperature and salinity variability in the subpolar North Atlantic over the past 10 k.y. [J]. Geology, 2007, 35(4): 315-318. doi: 10.1130/G23455A.1
[41] Clark P U, Dyke A S, Shakun J D, et al. The last glacial maximum [J]. Science, 2009, 325(5941): 710-714.
[42] Cortijo E, Labeyrie L, Vidal L, et al. Changes in sea surface hydrology associated with Heinrich event 4 in the North Atlantic Ocean between 40° and 60°N [J]. Earth and Planetary Science Letters, 1997, 146(1-2): 29-45. doi: 10.1016/S0012-821X(96)00217-8
[43] Bond G C, Lotti R. Iceberg discharges into the North Atlantic on millennial time scales during the last glaciation [J]. Science, 1995, 267(5200): 1005-1010. doi: 10.1126/science.267.5200.1005
[44] Xiao W S, Wang R J, Polyak L, et al. Stable oxygen and carbon isotopes in planktonic foraminifera Neogloboquadrina pachyderma in the Arctic Ocean: an overview of published and new surface-sediment data [J]. Marine Geology, 2014, 352: 397-408. doi: 10.1016/j.margeo.2014.03.024
[45] Missiaen L, Pichat S, Waelbroeck C, et al. Downcore variations of sedimentary detrital (238U/232Th) ratio: implications on the use of 230Thxs and 231Paxs to reconstruct sediment flux and ocean circulation [J]. Geochemistry, Geophysics, Geosystems, 2018, 19(8): 2560-2573. doi: 10.1029/2017GC007410
[46] Govin A, Braconnot P, Capron E, et al. Persistent influence of ice sheet melting on high northern latitude climate during the early Last Interglacial [J]. Climate of the Past, 2012, 8(2): 483-507. doi: 10.5194/cp-8-483-2012
[47] Zaric S, Donner B, Fischer G, et al. Sensitivity of planktic foraminifera to sea surface temperature and export production as derived from sediment trap data [J]. Marine Micropaleontology, 2005, 55(1-2): 75-105. doi: 10.1016/j.marmicro.2005.01.002
[48] Ottens J J. Planktic foraminifera as North Atlantic water mass indicators [J]. Oceanologica Acta, 1991, 14(2): 123-140.
[49] Morley A, Babila T L, Wright J, et al. Environmental controls on Mg/Ca in Neogloboquadrina incompta: A core-top study from the subpolar North Atlantic [J]. Geochemistry, Geophysics, Geosystems, 2017, 18(12): 4276-4298. doi: 10.1002/2017GC007111
[50] Irvali N, Galaasen E V, Ninnemann U S, et al. A low climate threshold for south Greenland Ice Sheet demise during the Late Pleistocene [J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(1): 190-195. doi: 10.1073/pnas.1911902116
[51] Villanueva J, Grimalt J O, Cortijo E, et al. Assessment of sea surface temperature variations in the central North Atlantic using the alkenone unsaturation index (U37k’) [J]. Geochimica et Cosmochimica Acta, 1998, 62(14): 2421-2427. doi: 10.1016/S0016-7037(98)00180-X
[52] Madureira L A S, Van Kreveld S A, Eglinton G, et al. Late Quaternary high-resolution biomarker and other sedimentary climate proxies in a Northeast Atlantic Core [J]. Paleoceanography and Paleoclimatology, 1997, 12(2): 255-269.
[53] Eynaud F, De Abreu L, Voelker A, et al. Position of the polar front along the western Iberian margin during key cold episodes of the last 45 ka [J]. Geochemistry, Geophysics, Geosystems, 2009, 10(7): Q07U05.
[54] Marchitto T M, Curry W B, Lynch-Stieglitz J, et al. Improved oxygen isotope temperature calibrations for cosmopolitan benthic foraminifera [J]. Geochimica et Cosmochimica Acta, 2014, 130: 1-11. doi: 10.1016/j.gca.2013.12.034
[55] Curry W B, Oppo D W. Glacial water mass geometry and the distribution of δ13C of ΣCO2 in the western Atlantic ocean [J]. Paleoceanography and Paleoclimatology, 2005, 20(1): PA1017.
[56] Keigwin L D, Boyle E A. Late quaternary paleochemistry of high-latitude surface waters [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1989, 73(1-2): 85-106. doi: 10.1016/0031-0182(89)90047-3
[57] Mook W G, Bommerson J C, Staverman W H. Carbon isotope fractionation between dissolved bicarbonate and gaseous carbon dioxide [J]. Earth and Planetary Science Letters, 1974, 22(2): 169-176. doi: 10.1016/0012-821X(74)90078-8
[58] Zhan R, Winn K, Sarnthein M. Benthic foraminiferal δ13C and accumulation rates of organic carbon: Uvigerina Peregrina group and Cibicidoides Wuellerstorfi [J]. Paleoceanography and Paleoclimatology, 1986, 1(1): 27-42.
[59] Lynch-Stieglitz J, Fairbanks R G, Charles C D. Glacial-interglacial history of Antarctic intermediate water: relative strengths of Antarctic versus Indian Ocean sources [J]. Paleoceanography and Paleoclimatology, 1994, 9(1): 7-29.
[60] Polyak L, Curry W B, Darby D A, et al. Contrasting glacial/interglacial regimes in the western Arctic Ocean as exemplified by a sedimentary record from the Mendeleev Ridge [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2004, 203(1-2): 73-93. doi: 10.1016/S0031-0182(03)00661-8
[61] 李铁刚, 孙荣涛, 张德玉, 等. 晚第四纪对马暖流的演化和变动: 浮游有孔虫和氧碳同位素证据[J]. 中国科学 D辑: 地球科学, 2007, 50(5):725-735 doi: 10.1007/s11430-007-0003-2 LI Tiegang, SUN Rongtao, ZHANG Deyu, et al. Evolution and variation of the Tsushima warm current during the late quaternary: Evidence from planktonic foraminifera, oxygen and carbon isotopes [J]. Science in China Series D: Earth Sciences, 2007, 50(5): 725-735. doi: 10.1007/s11430-007-0003-2
[62] Elderfield H, Vautravers M, Cooper M. The relationship between shell size and Mg/Ca, Sr/Ca, δ18O, and δ13C of species of planktonic foraminifera [J]. Geochemistry, Geophysics, Geosystems, 2002, 3(8): 1-13.
[63] Donner B, Wefer G. Flux and stable isotope composition of Neogloboquadrina pachyderma and other planktonic foraminifers in the southern ocean (Atlantic sector) [J]. Deep Sea Research Part I: Oceanographic Research Papers, 1994, 41(11-12): 1733-1743. doi: 10.1016/0967-0637(94)90070-1

计量
- 文章访问数: 7235
- HTML全文浏览量: 594
- PDF下载量: 88


