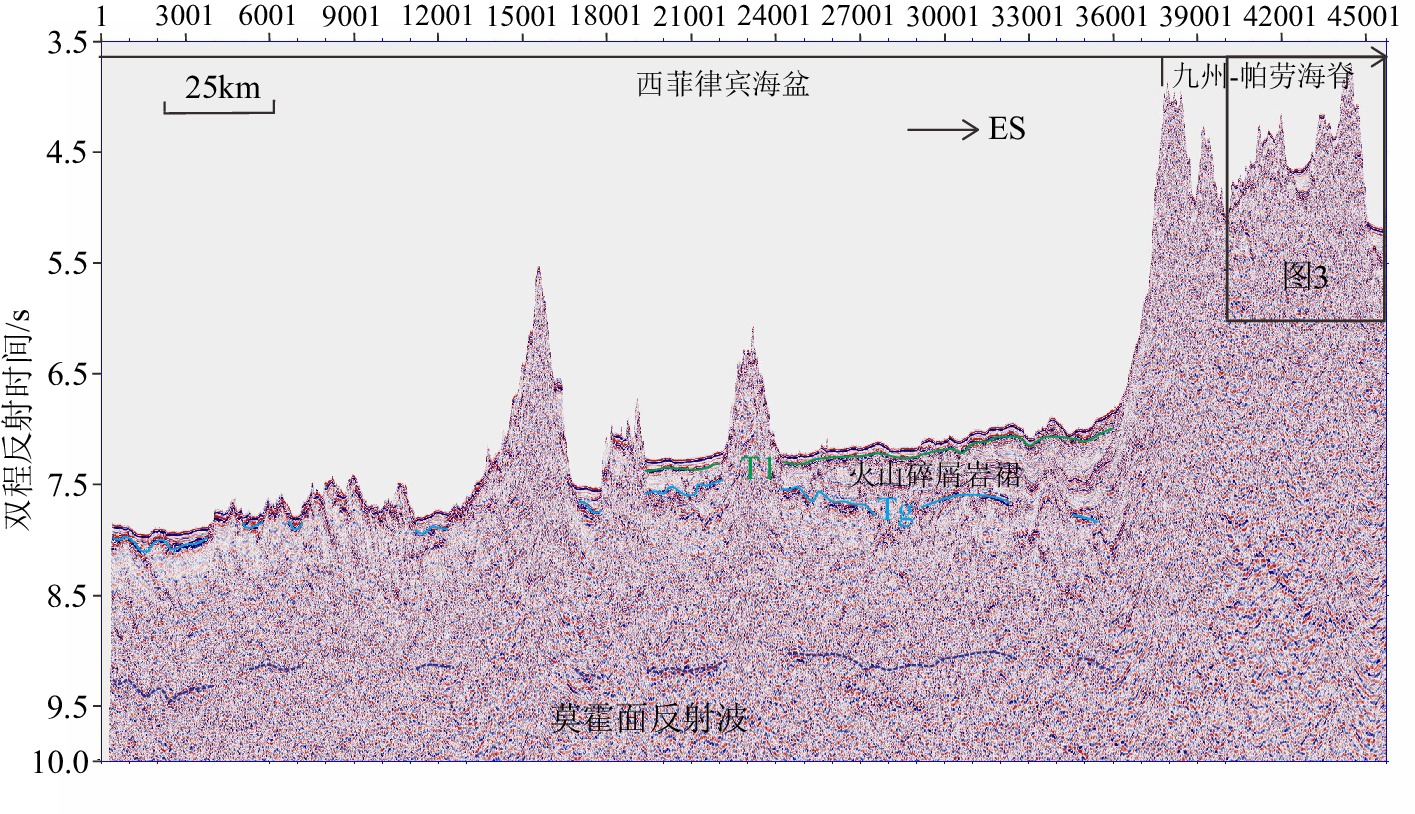
Tectono-sedimentary characteristics and subduction initiation in the middle Kyushu-Palau Ridge and adjacent basins: A comprehensive study of multichannel seismic reflection profiles
-
摘要: 九州-帕劳海脊及两侧的盆地是研究边缘海形成演化和板块俯冲起始机制的理想区域。利用最新采集的深反射地震数据并结合钻孔数据,研究了九州-帕劳海脊及其东西两侧盆地的构造沉积特征及地壳结构,探讨了西菲律宾海的起源及沿古伊豆-博宁-马里亚纳弧的俯冲起始机制。地震剖面和钻孔岩心揭示西菲律宾海盆发育上、下两套不同来源的沉积物盖层,其中下部地层厚度差别大,多为火山物质,来源于九州-帕劳海脊的岛弧火山作用,表现为向海脊方向的持续增厚,靠近海脊的山脚下发育火山碎屑岩裙;上部地层厚度较为稳定,为一套大洋的深水细粒沉积物。西菲律宾海盆和帕里西维拉海盆的地壳厚度约为6~8 km,接近全球平均洋壳厚度;西菲律宾海盆的莫霍面呈宽缓的褶皱状,与沉积物之下的洋壳基底基本同步起伏。西菲律宾海盆在中生代可能位于南半球澳大利亚的北缘,是在陆缘弧(包含部分大陆碎片)的基础上,因古近纪弧间伸展和海底扩张而逐步发展起来的。西菲律宾海盆的地震剖面和钻井均揭示了始新世时期的构造挤压事件,沿古伊豆-博宁-马里亚纳弧的俯冲起始可能是印亚碰撞的远场效应引起的诱导俯冲,俯冲过程伴随着侧向传播和持续的挤压应力场,直到约30 Ma开始岛弧裂离。该研究对揭示东南亚板块构造演化过程和板块俯冲起始机制具有一定的理论意义。Abstract: The Kyushu Palau Ridge and the basins on the both sides are ideal areas for understanding the formation and evolution of marginal seas and the initiation mechanism of plate subduction. We studied the tectonic sedimentary characteristics and crustal structure of the Kyushu-Palau Ridge (KPR) and adjacent basins using newly collected deep reflection seismic data combined with the borehole data, and discussed the origin and subduction initiation mechanism of the West Philippines Sea. Seismic profiles and drill cores revealed that two sets of sediment caprocks are developed from different sources in the Western Philippines Basin. The thickness of the lower set is very different, and most of them are volcanic materials that derived from island arc volcanos of KPR, showing continuous thickening towards KPR, and volcanoclastic rock skirts are developed near the foot of KPR. The thickness of the upper set is relatively stable, being composed of deep-water fine-grained sediments. The crust thickness of the western Philippines Basin and Parece Vela Basin is 6-8km, which is similar to the global average oceanic crust thickness. The Moho surface of the Western Philippines Basin is in a wide-gentle fold shape, and fluctuates synchronously with the ocean crust basement beneath sediments. The Western Philippines Basin might be located in the northern margin of Australia in the southern hemisphere in the Mesozoic, and was gradually developed on a continental margin arc (including some continental fragments) due to inter-arc extension and submarine spreading in the Paleogene. Seismic profiles and drilling in the Western Philippines Basin have revealed tectonic compression events during the Eocene. The subduction along the paleo-IBM (Izu-Bonin-Mariana) might be induced by the remote effect of the Indo-Asian collision. The subduction process was accompanied by lateral propagation and persistent compressive stress field, which consequently resulted in the island arc broke up in about 30Ma. This study has certain theoretical significance for studying the plate tectonic evolution and subduction initiation mechanism in Southeast Asia.
-
作为一种古海洋代用指标,浮游有孔虫壳体重量具体是指限定粒径范围内浮游有孔虫单种一定数量壳体的单枚平均重量,可代表样品中浮游有孔虫壳体的壳壁厚度。该指标首先由Lohmann[1]提出,其发现特定海区内表层沉积物样品中浮游有孔虫壳体重量随着样品水深的增加而减小,认为壳体重量实际上反映了深海碳酸钙溶解作用,即浮游有孔虫壳体受到溶解作用影响后壳壁变薄,表现为壳体重量变轻。从原理上,深海碳酸钙溶解作用受海水碳酸钙饱和程度(Ω = [Ca2+][CO32-]/K*sp,K*sp为溶解常数)的控制,理论上当Ω<1时溶解作用发生[2]。由于现代海水[Ca2+]基本保持恒定[3],所以Ω可近似= [CO32-]/K*sp,因此,浮游有孔虫壳体重量可作为一种潜在的深海[CO32-]指标。海水[CO32-]与溶解无机碳含量(DIC)、碱度(ALK)等海洋碳酸盐系统参数相关,海水[CO32-]重建是目前量化海洋碳循环过程演化的主要手段之一[4]。
Broecker等[5]首次尝试建立浮游有孔虫壳体重量与深海[CO32-]的经验关系,通过对热带大洋表层沉积物样品进行研究,发现浮游有孔虫Pulleniatina obliquiloculata、Neogloboquadrina dutertrei和Trilobatus sacculifer壳体重量与当地深海碳酸根离子饱和程度(Δ[CO32-] = [CO32-] - [CO32-]/Ω)呈线性正相关,确证了浮游有孔虫壳体重量可作为一种深海[CO32-]代用指标。但是,上述现代过程校准结果也存在如下问题:在Δ[CO32-]高值时,壳体重量与其相关性较差;不同海区之间壳体重量与Δ[CO32-]的关系有偏差。这意味着沉积物中浮游有孔虫壳体重量还应受到其他因素的影响。
随后,另一项现代过程校准工作指出了上述问题的症结所在[6]。北大西洋表层沉积物中浮游有孔虫壳体重量与表层海水温度、碳酸盐系统参数存在显著相关性,认为此时壳体重量反映了浮游有孔虫在上层海洋生长过程中的钙化程度。海洋生物的钙化过程指浮游有孔虫、颗石藻等钙化生物根据方程Ca2++2HCO3-→CaCO3+CO2+H2O,形成 CaCO3壳体,钙化程度强弱也控制着浮游有孔虫壳体的壳壁厚度(即壳体重量)。于是,浮游有孔虫壳体重量开始被用作一种指示钙化程度的指标[7-8]。但是,在这一思路下利用该指标进行长时间尺度古海洋重建时又遇到了问题。现代北大西洋深海水团年龄较轻(含更少的呼吸CO2),其深海碳酸钙溶解作用较弱,表层沉积物中浮游有孔虫壳体重量更多反映其钙化程度。但是在地质历史中,碳酸钙溶解作用有可能剧烈下降,比如在约200~600 ka的中布容溶解事件中,北大西洋沉积物柱状样中Globigerina bulloides壳体重量明显下降,事件中壳体重量的变化实际上更多反映了深海溶解作用变化,而非浮游有孔虫的钙化程度[9]。
无法区分沉积物中浮游有孔虫壳体重量中包含的钙化信息以及溶解信息,是限制该指标古海洋学应用的主要原因。浮游有孔虫在上层海洋生长钙化,死亡后沉降至深海沉积物中埋藏,其壳体重量中钙化信息即浮游有孔虫死亡沉降前的初始壳体重量,而溶解信息即初始壳体重量受到深海溶解作用后的减轻部分。浮游有孔虫壳体重量可作为一种古海洋指标(钙化程度或深海Δ[CO32-])的前提在于保证其在指定时间尺度下只受到初始壳体重量变化或深海溶解作用单一因素的影响。这需要回答以下核心科学问题:浮游有孔虫壳体开始受到溶解作用影响的环境条件是什么?浮游有孔虫壳体重量对钙化过程、溶解过程的响应敏感度是怎样的?
1. 基于全球热带大洋表层沉积物的现代过程校准
1.1 P. obliquiloculata壳体重量受控钙化作用和溶解作用
基于表层沉积物的现代过程校准是建立古海洋指标的核心基础。目前,在中低纬海洋,互相之间可类比的能兼顾不同水深的表层沉积物浮游有孔虫壳体重量资料主要来自全球热带大洋——翁通爪哇海台(Ontong Java Plateau)[5, 10]、帕劳海脊(Palau Ridge)[10]、东经九十度海脊(Ninetyeast Ridge)和塞阿拉高地(Ceara Rise)[5] (图1A),涉及的浮游有孔虫属种包括P. obliquiloculata、N. dutertrei和T. sacculifer,壳体的粒径范围为355~400 μm。首先来看P. obliquiloculata,毫无疑问,在全球范围内,其壳体重量与深海Δ[CO32-]存在线性关系,但是这一相关性(r = 0.52)并不高(图1B)。也就是说深海溶解作用只是控制P. obliquiloculata壳体重量的因素之一。细化到单一海区,翁通爪哇海台、帕劳海脊和东经九十度海脊区P. obliquiloculata壳体重量与深海Δ[CO32-]高度正相关(图1C),表明在上述单一海域,深海Δ[CO32-] / 溶解作用是控制表层沉积物中P. obliquiloculata壳体重量的唯一主导因素。而在大西洋塞阿拉高地,整体上壳体重量与深海Δ[CO32-]相关性并不好,原因在于大西洋深海的弱溶解作用(高Δ[CO32-]值)。当剔除深海Δ[CO32-] >20 μmol·kg−1的站位后,塞阿拉高地P. obliquiloculata壳体重量就和Δ[CO32-]呈现了明显正相关性(图1C)。
![图 1 全球热带大洋表层沉积物P. obliquiloculata壳体重量与深海Δ[CO32-]的关系[14]]() 图 1 全球热带大洋表层沉积物P. obliquiloculata壳体重量与深海Δ[CO32-]的关系[14]A: 表层沉积物站位图,B–C: 全球热带大洋以及单一海区中P. obliquiloculata壳体重量与深海Δ[CO32-]的关系。图C中阴影部分代表线性拟合的95 %置信区间。Figure 1. Shell weights of P. obliquiloculata from global tropical surface-sediment samples versus deep-ocean Δ[CO32-] [14]A: sites of surface-sediment sampling, B-C: relationships between P. obliquiloculata shell weight and deep-ocean Δ[CO32-] in the global tropical oceans (B) and in specific regions (C). Shaded areas in (C) denote 95% confidence intervals.
图 1 全球热带大洋表层沉积物P. obliquiloculata壳体重量与深海Δ[CO32-]的关系[14]A: 表层沉积物站位图,B–C: 全球热带大洋以及单一海区中P. obliquiloculata壳体重量与深海Δ[CO32-]的关系。图C中阴影部分代表线性拟合的95 %置信区间。Figure 1. Shell weights of P. obliquiloculata from global tropical surface-sediment samples versus deep-ocean Δ[CO32-] [14]A: sites of surface-sediment sampling, B-C: relationships between P. obliquiloculata shell weight and deep-ocean Δ[CO32-] in the global tropical oceans (B) and in specific regions (C). Shaded areas in (C) denote 95% confidence intervals.全球范围内P. obliquiloculata壳体重量与深海Δ[CO32-]的关系提供了以下关键信息。首先,Δ[CO32-] = 20 μmol·kg−1似乎是深海溶解作用开始影响浮游有孔虫壳体的阈值,与前人基于培养实验得出的结论一致[11-12]。理论上来说,海洋中碳酸钙质壳体应该在Ω<1(即Δ[CO32-]<0 μmol·kg−1)时才会开始受到溶解作用的影响。但是实际上,这一Δ[CO32-]阈值高达20 μmol·kg−1,其中的原因应在于浮游有孔虫壳体实际上不是纯CaCO3质,含有其他元素(特别是Mg)会导致其更容易受到溶解作用的影响[13]。另外,样品涉及四个海区内P. obliquiloculata壳体重量与深海Δ[CO32-]的线性拟合方程体现了基本一致的斜率值(Δ[CO32-]每降低1 μmol·kg−1时壳体重量的下降值),只不过线性拟合方程的截距有显著差别。具体来说,当固定深海Δ[CO32-]值时,4个海区P. obliquiloculata壳体重量有着明显差别,从重到轻依次为翁通爪哇海台、帕劳海脊、塞阿拉高地和东经九十度海脊。以上说明,初始壳体重量不同是导致4个海区之间壳体重量-深海Δ[CO32-]关系差别的主要原因,而且,P. obliquiloculata初始壳体重量信息在受到溶解作用影响后依然能完好保留下来。简而言之,沉积物中P. obliquiloculata壳体重量应同时包含钙化作用信息和溶解作用信息。
1.2 N. dutertrei壳体重量主要受控溶解作用
全球范围内N. dutertrei壳体重量与深海Δ[CO32-]同样存在显著的线性关系,不过其相关性(r=0.88)明显高于P. obliquiloculata的情况(r=0.52)(图2A)。类似的,单一海区内,N. dutertrei壳体重量与深海Δ[CO32-]的线性拟合方程体现了基本一致的斜率值,相比于P. obliquiloculata,线性拟合方程的截距差距较小。也就是说,对比P. obliquiloculata,沉积物中N. dutertrei壳体重量对深海Δ[CO32-] / 溶解作用的敏感度更高,原因应在于P. obliquiloculata的抗溶性高于N. dutertrei [15]。简而言之,沉积物中N. dutertrei壳体重量主要受到深海Δ[CO32-] / 溶解作用的控制,相对来说钙化作用控制的初始壳体重量变化对其影响较小,N. dutertrei壳体重量可作为一种潜在的深海Δ[CO32-] / 溶解作用代用指标。
![图 2 全球热带大洋(A)以及单一海区中(B)表层沉积物N. dutertrei壳体重量与深海Δ[CO32-]的关系[14]]() 图 2 全球热带大洋(A)以及单一海区中(B)表层沉积物N. dutertrei壳体重量与深海Δ[CO32-]的关系[14]图B中阴影部分代表线性拟合的95 %置信区间。Figure 2. Relationships between shell weights of N. dutertrei in surface-sediment samples from global tropical (A) or specific regions (B) and deep-ocean Δ[CO32-] [14]Shaded areas in (B) denote 95% confidence intervals.
图 2 全球热带大洋(A)以及单一海区中(B)表层沉积物N. dutertrei壳体重量与深海Δ[CO32-]的关系[14]图B中阴影部分代表线性拟合的95 %置信区间。Figure 2. Relationships between shell weights of N. dutertrei in surface-sediment samples from global tropical (A) or specific regions (B) and deep-ocean Δ[CO32-] [14]Shaded areas in (B) denote 95% confidence intervals.1.3 T.sacculifer壳体重量受控溶解作用
T. sacculifer是典型的易溶种,其抗溶性远弱于P. obliquiloculata和N. dutertrei[15]。不出意料,T. sacculifer壳体重量与深海Δ[CO32-]的相关性最好(剔除深海Δ[CO32-]>20 μmol·kg−1的站位后)(图3)。单一海区内,T. sacculifer壳体重量与深海Δ[CO32-]的线性拟合方程体现了基本一致的斜率值,更重要的是,四个海区的线性拟合方程的截距几乎没有差别(图3B)。这四个海区不同的上层海洋环境必定导致T. sacculifer的初始壳体重量不同,证据一是这些海区中P. obliquiloculata和N. dutertrei的初始壳体重量就存在显著差距(图1-2),证据二则是当Δ[CO32-]>20 μmol·kg−1时(此时壳体重量等同于初始壳体重量),T. sacculifer的(初始)壳体重量存在明显的变化幅度(图3)。但是,一旦T. sacculifer壳体开始受到溶解作用(Δ[CO32-]<20 μmol·kg−1),其初始壳体重量信号几乎被迅速抹除。也就是说,易溶种T. sacculifer的壳体对深海Δ[CO32-] / 溶解作用极其敏感,在深海Δ[CO32-]<20 μmol·kg−1的情况下,其壳体重量基本只反映溶解信号。也就是说,T. sacculifer壳体重量是一种绝佳的深海Δ[CO32-] / 溶解作用代用指标。
![图 3 全球热带大洋(A)及单一海区中(B)表层沉积物T. sacculifer壳体重量与深海Δ[CO32-]的关系]() 图 3 全球热带大洋(A)及单一海区中(B)表层沉积物T. sacculifer壳体重量与深海Δ[CO32-]的关系图B中阴影部分代表线性拟合的95%置信区间。Figure 3. Relationships between shell weights of T. sacculifer from global tropical (A) surface-sediment or specific regions (B) and deep-ocean Δ[CO32-] [14]Shaded areas in (B) denote 95% confidence intervals.
图 3 全球热带大洋(A)及单一海区中(B)表层沉积物T. sacculifer壳体重量与深海Δ[CO32-]的关系图B中阴影部分代表线性拟合的95%置信区间。Figure 3. Relationships between shell weights of T. sacculifer from global tropical (A) surface-sediment or specific regions (B) and deep-ocean Δ[CO32-] [14]Shaded areas in (B) denote 95% confidence intervals.2. 基于南大洋南极带表层沉积物的现代过程校准
浮游有孔虫Neogloboquadrina pachyderma
(sinistral)是一种典型的中高纬度种,在南大洋是当地的绝对优势种[16]。基于阿蒙森海(Amundsen Sea)、罗斯海(Ross Sea)和普里兹湾(Prydz Bay)表层沉积物的现代过程校准工作评估了南大洋南极带浮游有孔虫N. pachyderma(sin.)壳体重量指标的环境指示意义[17]。出乎意料的是,校准结果与前述热带大洋浮游有孔虫P. obliquiloculata、N. dutertrei和T. sacculifer的情况并不完全一致。当站位深海Δ[CO32-]>20 μmol·kg−1时, 三个海区N. pachyderma(sin.)壳体重量变化幅度较大(图4),表明其记录了复杂的上层海洋钙化作用信号,这和热带大洋的研究结果一致。但是,当站位深海Δ[CO32-]<20 μmol·kg−1时,除了300~355 μm壳体粒径范围以外,N. pachyderma(sin.)壳体重量与深海Δ[CO32-]并无显著相关性。甚至在300~355 μm壳体粒径范围内,由于统计范围内只包含3个数据点,其形成的壳体重量-Δ[CO32-]线性拟合关系也并不十分可靠。 ![图 4 南大洋南极带表层沉积物N. pachyderma(sin.)壳体重量与深海Δ[CO32-]的关系[17]]() 图 4 南大洋南极带表层沉积物N. pachyderma(sin.)壳体重量与深海Δ[CO32-]的关系[17]A:表层沉积物站位图,B-D:分别为3种不同粒径范围200~250、250~300和300~355 μm的结果。Figure 4. Relationships between shell weights of N. pachyderma (sin.) from Antarctic Zone surface-sediment samples and deep-ocean Δ[CO32-] [17]A: sites of surface-sediment samples; B–D: results for three different shell size ranges 200~250 μm (B), 250~300 μm (C), and 300~355 μm (D)
图 4 南大洋南极带表层沉积物N. pachyderma(sin.)壳体重量与深海Δ[CO32-]的关系[17]A:表层沉积物站位图,B-D:分别为3种不同粒径范围200~250、250~300和300~355 μm的结果。Figure 4. Relationships between shell weights of N. pachyderma (sin.) from Antarctic Zone surface-sediment samples and deep-ocean Δ[CO32-] [17]A: sites of surface-sediment samples; B–D: results for three different shell size ranges 200~250 μm (B), 250~300 μm (C), and 300~355 μm (D)由此可以推断,南大洋南极带沉积物中N. pachyderma(sin.)壳体重量主要显示了上层海洋的钙化过程信号,而非深海的碳酸钙溶解作用。其中的原因可能是:相比于热带大洋,高纬海洋环境并不适宜浮游有孔虫的生长钙化,具体表现为高纬海洋浮游有孔虫的数量以及分异度都显著低于中低纬,在这种极端环境下,浮游有孔虫对上层海洋生长环境变化更加敏感;另外,南极带极端海洋环境下N. pachyderma(sin.)初始壳体比较脆弱(壳壁薄、重量轻),当其受到溶解作用影响后极易破碎,这导致沉积物中剩余的完整壳体可能并不具备整体代表性(只有少数重量偏重的壳体能保存下来),因此,壳体重量与深海溶解作用的相关性不高。以上可能共同导致南大洋南极带沉积物中N. pachyderma
(sin.)壳体重量主要反映初始壳体重量,因此,可作为钙化程度指标。 3. 浮游有孔虫壳体重量的古海洋应用模式
基于上述现代过程校准结果:深海溶解作用开始影响沉积物中浮游有孔虫壳体重量的阈值为Δ[CO32-] = 20 μmol·kg−1,因此当地质历史中研究站位的深海Δ[CO32-]始终>20 μmol·kg−1时,壳体重量可作为浮游有孔虫钙化程度指标;在受到溶解作用影响后(Δ[CO32-]<20 μmol·kg−1),不同浮游有孔虫种壳体重量对Δ[CO32-]响应敏感度并不一致,相对来说,易溶种,比如T. sacculifer,其壳体重量更适合作为深海Δ[CO32-]代用指标。
3.1 浮游有孔虫壳体重量反映钙化程度
浮游有孔虫、颗石藻等海洋生物的钙化过程以1∶2的比例消耗上层海水中的DIC和ALK,总体将导致上层海水pH下降、pCO2上升[2],因此该过程在海洋碳循环领域具有重要的研究意义。目前使用浮游有孔虫壳体重量作为钙化程度指标的研究多位于大西洋,因为大西洋深海的Δ[CO32-]较高、碳酸钙溶解作用较弱。因此大西洋基于壳体重量的古海洋研究多探讨浮游有孔虫钙化程度的受控机制。
末次冰期以来北大西洋浮游有孔虫G. bulloides
壳体重量随着大气pCO2升高/海洋酸化而下降,意味着该种的钙化过程可能受控海水碳酸盐系统[6]。这一结论得到了一系列培养实验和现代过程调查的支持[7, 18-22],这些研究均发现特定浮游有孔虫钙化程度变化与其生活水深的海水pH/[CO32-]正相关,因此在海-气CO2平衡海区,大气pCO2升高引发的上层海洋酸化会导致浮游有孔虫钙化程度减弱,反映为壳体重量下降。然而,随着在全球各海区针对不同浮游有孔虫种开展研究,逐渐认识到无论是在现代还是地质历史中,浮游有孔虫钙化过程机制远比我们想象的复杂,除了海水碳酸盐系统,海水温度[14, 17, 23]、盐度[24-25]以及营养盐浓度[26]等其他环境参数也是潜在的影响因素,其中温度的影响甚至被认为和海水碳酸盐系统的影响相当[27]。比如,利用深海Δ[CO32-]校正得到的P. obliquiloculata初始壳体重量结果显示无论在现代还是地质历史中,P. obliquiloculata钙化程度变化均与上层海水温度显著正相关,而和海水碳酸盐系统参数并无关联[14]。类似地,在南大洋南极带,表层沉积物中N. pachyderma(sin.)的初始壳体重量也与现代上层海水温度呈正相关[17]。这些研究指出,至少对于P. obliquiloculata和N. pachyderma(sin.),其钙化程度主要受控于海水温度而非海洋酸化。以上可以看出,浮游有孔虫钙化作用对海洋环境要素的响应方式应存在明显的种间差异。 3.2 浮游有孔虫壳体重量重建深海碳酸盐系统
海水碳酸盐系统中[CO32-]反映了海水ALK和DIC的变化,ALK代表海水碳酸盐系统的缓冲能力,DIC则代表当前海水储碳量状况。目前,古深海[CO32-]指标是探索海洋碳储库演化的主要手段之一。正如前文(1.2节和1.3节)所述,当深海Δ[CO32-]<20 μmol·kg−1时,浮游有孔虫N. dutertrei和T. sacculifer壳体重量可作为可靠的Δ[CO32-]代用指标。于是,Qin等[28]基于全球热带大洋表层沉积物样品建立了N. dutertrei和T. sacculifer的壳体重量-深海Δ[CO32-]经验校准公式(图5)。
![图 5 T. sacculifer和N. dutertrei的壳体重量-深海Δ[CO32-]的经验校准公式[28]]()
正是通过N. dutertrei和T. sacculifer壳体重量,重建了热带西太平洋上新世以来不同时间尺度上深海Δ[CO32-]变化[10, 28-30]。热带太平洋适合开展该指标古海洋学应用的原因在于:该海区沉积物中含有丰富的浮游有孔虫;另外太平洋的深海Δ[CO32-]足够低,至少可以保证上新世以来Δ[CO32-]始终显著低于20 μmol·kg−1。基于Δ[CO32-] ≈ ALK – DIC的关系[31],深海Δ[CO32-]记录可用来探讨地质历史中深海碳储库/碳循环演化过程。热带西太平洋这一系列深海Δ[CO32-]记录揭示了如下结论:冰期旋回中太平洋深海碳酸盐系统通过碳酸钙补偿机制响应海平面控制的陆架珊瑚礁碳酸钙埋藏量变化[10, 32];量化了中布容溶解事件中太平洋深海Δ[CO32-]的下降幅度[29];中更新世气候转型期间太平洋深海DIC储库显著增强[28, 33];南极冰盖/海冰扩张有利于太平洋深海储碳,而AMOC主要调节大西洋和太平洋深海的DIC分配模式[30]等。
4. 浮游有孔虫壳体重量指标的优劣势以及未来展望
当用来反映钙化程度,浮游有孔虫壳体重量指标的应用前提为保证样品始终不受到溶解作用的影响,即需要深海Δ[CO32-]>20 μmol·kg−1。图1可以看到,即使对于典型抗溶种P. obliquiloculata,一旦Δ[CO32-]<20 μmol·kg−1,其壳体重量会开始显著下降。因此,在指标使用中,可通过底栖有孔虫B/Ca比值等手段恢复研究时段内深海Δ[CO32-]变化;或者基于站位现代深海Δ[CO32-]数据以及预估研究时段中Δ[CO32-]的可能变化幅度,来校准或排除溶解作用的干扰。
目前成熟的深海Δ[CO32-]代用指标主要包括特定浮游有孔虫壳体重量和底栖有孔虫B/Ca比值[31],两者均基于全球大洋表层沉积物建立的经验校准公式。底栖有孔虫B/Ca比值指标的优势在于其Δ[CO32-]适用范围高达−20~80 μmol·kg−1,该指标几乎可应用于所有海区,特别是Δ[CO32-]普遍较高的大西洋。相比之下,浮游有孔虫壳体重量指标的Δ[CO32-]适用范围只有−20~20 μmol·kg−1,目前来看,该指标适合于全球平均深海Δ[CO32-]最低的太平洋,以及其他海域水深较深的站位。壳体重量指标的优势则在于浮游有孔虫(相较于底栖有孔虫)的易获取性,这意味着在有孔虫含量丰富的站位,该指标可形成高分辨率的重建记录。
以上关于浮游有孔虫壳体重量指标的认识主要来自于表层沉积物的现代过程校准结果,未来建议进一步加强基于现场观测、实验室培养等方式的浮游有孔虫生长钙化和溶解过程机制研究。比如,浮游有孔虫在生长钙化过程中会在壳体内以及周围形成一个独特的“微环境”,并能通过生命效应以提高微环境中pH以及[CO32-]来促进生长钙化[34],这种生命效应过程目前对学界而言仍是一个“黑盒子”;浮游有孔虫在不同生长阶段的钙化过程对海洋环境的响应方式可能存在显著差异,处于成长期(小尺寸)时由于只能吸收有限体积海水中的离子进行钙化,其钙化作用反而会在高DIC浓度(低pH值)环境下增强[21];现代观测研究指出相当一部分上层海洋生产的CaCO3会在浅水透光层被溶解,其机制可能与透光层大量有机碳降解导致的海洋酸化有关[35-36],而这一特殊溶解机制对沉积物中浮游有孔虫壳体的影响尚不清楚。上述这些细节问题无疑需要更加精细的观测与培养研究来解决。
5. 总结
基于目前已有的全球大洋表层沉积物数据,综述了浮游有孔虫壳体重量作为钙化程度或者深海Δ[CO32-]指标的现代过程校准结果。深海溶解作用开始影响浮游有孔虫壳体的阈值是Δ[CO32-]=20 μmol·kg−1,在此阈值以上,壳体重量可用来反映浮游有孔虫钙化程度变化。在此阈值以下,浮游有孔虫易溶种T. sacculifer和N. dutertrei的壳体重量可作为深海Δ[CO32-]代用指标。目前基于浮游有孔虫壳体重量指标的古海洋研究显示:在钙化过程受控机制方面,海水碳酸盐系统和温度是影响浮游有孔虫钙化作用的主导因素,不同浮游有孔虫种钙化过程对海洋环境要素的响应方式存在明显差异;在深海Δ[CO32-]重建方面,海平面、南大洋冰盖/海冰以及全球温盐环流变化是控制上新世以来太平洋深海碳循环的主导因素。
-
![]()
图 1 菲律宾海构造简图
底图由Global Mapper软件制作,高程数据来自http://www.ngdc.noaa.gov/mgg/global/global.html,磁条带数据据文献[12]。
Figure 1. Tectonic setting of the Philippine Sea
The base map was drawn by Global Mapper software and elevation data was from http://www.ngdc.noaa.gov/mgg /global/global.html. Magnetic bands data are from reference [12].
![]()
图 2 DSDP59-447井层位标定图(MCS01线)
Figure 2. Seismic interpretation in stratigraphy and structural geology for Well DSDP59-447 (Line MCS01)
![]()
图 4 DSDP59-449井层位标定图(MCS02线)
Figure 4. Seismic interpretation in stratigraphy for Well DSDP59-449 (Line MCS02)
![]()
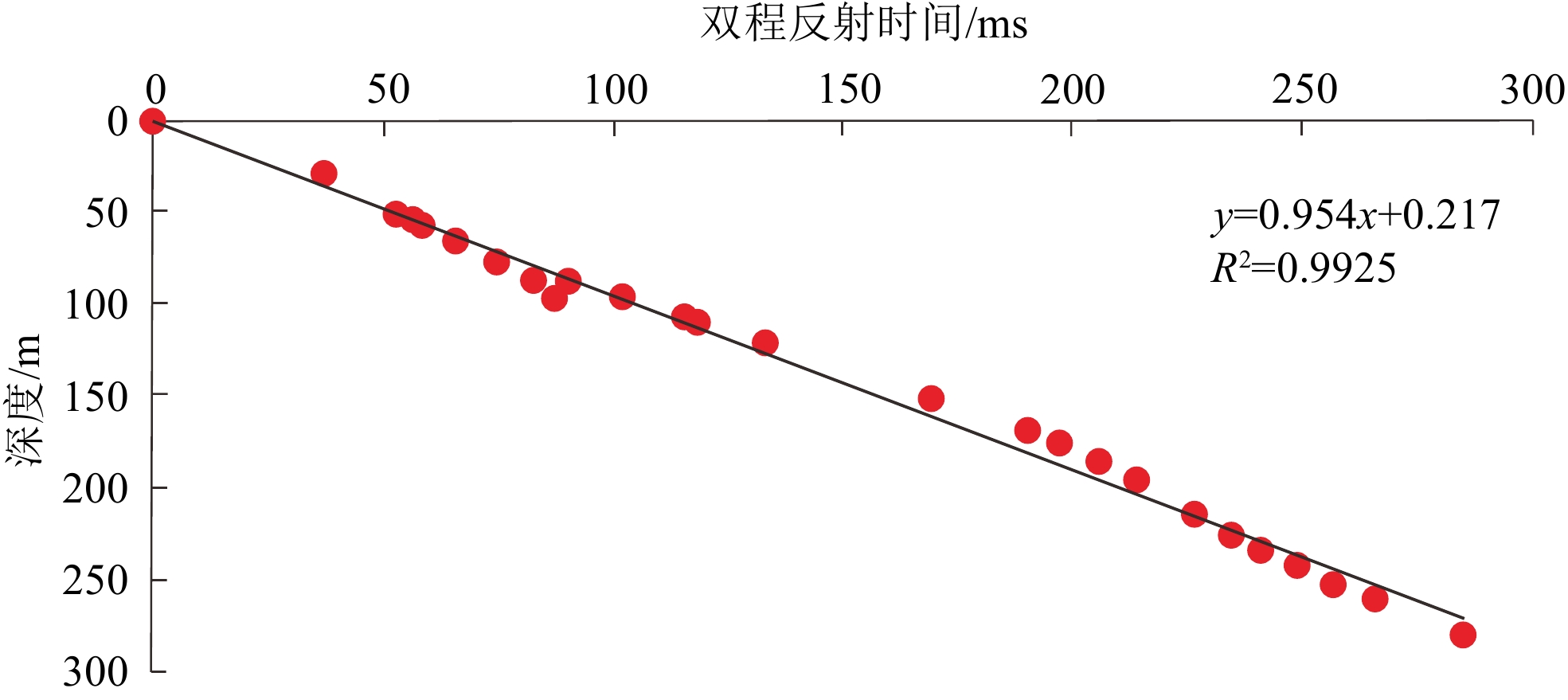
图 5 DSDP59-447、449井时深转换拟合图
Figure 5. Time-depth conversion fitting of Wells DSDP59-447 and 449
![]()
图 3 DSDP59-448井层位标定图(MCS03线)
Figure 3. Seismic interpretation in stratigraphy and structural geology for Well DSDP59-448 (Line MCS03)
-
[1] Seno T, Maruyama S. Paleogeographic reconstruction and origin of the Philippine Sea [J]. Tectonophysics, 1984, 102(1-4): 53-84. doi: 10.1016/0040-1951(84)90008-8
[2] Hall R, Ali J R, Anderson C D, et al. Origin and motion history of the Philippine Sea Plate [J]. Tectonophysics, 1995, 251(1-4): 229-250. doi: 10.1016/0040-1951(95)00038-0
[3] Hall R. Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, model and animations [J]. Journal of Asian Earth Sciences, 2002, 20(4): 353-431. doi: 10.1016/S1367-9120(01)00069-4
[4] Pubellier M, Ali J, Monnier C. Cenozoic Plate interaction of the Australia and Philippine Sea Plates: “hit-and-run” tectonics [J]. Tectonophysics, 2003, 363(3-4): 181-199. doi: 10.1016/S0040-1951(02)00671-6
[5] Yamazaki T, Takahashi M, Iryu Y, et al. Philippine Sea Plate motion since the Eocene estimated from paleomagnetism of seafloor drill cores and gravity cores [J]. Earth, Planets and Space, 2010, 62(6): 495-502. doi: 10.5047/eps.2010.04.001
[6] Wu J, Suppe J, Lu R Q, et al. Philippine Sea and East Asian plate tectonics since 52Ma constrained by new subducted slab reconstruction methods [J]. Journal of Geophysical Research:Solid Earth, 2016, 121(6): 4670-4741. doi: 10.1002/2016JB012923
[7] Arculus R J, Ishizuka O, Bogus K A, et al. A record of spontaneous subduction initiation in the Izu-Bonin-Mariana arc [J]. Nature Geoscience, 2015, 8(9): 728-733. doi: 10.1038/ngeo2515
[8] Stern R J, Gerya T. Subduction initiation in nature and models: a review [J]. Tectonophysics, 2018, 746: 173-198. doi: 10.1016/j.tecto.2017.10.014
[9] 丁巍伟, 李家彪. 九州-帕劳海脊南段的深部结构探测及对板块俯冲起始机制的可能启示[J]. 海洋地质与第四纪地质, 2019, 39(5):98-103 DING Weiwei, LI Jiabiao. Seismic detection of deep structure for Southern Kyueshu-Palau Ridge and its possible implications for subduction initiation [J]. Marine Geology & Quaternary Geology, 2019, 39(5): 98-103.
[10] Sun W D, Zhang L P, Li H, et al. The synchronic Cenozoic subduction initiations in the west Pacific induced by the closure of the neo-Tethys ocean [J]. Science Bulletin, 2020, 65(24): 2068-2071. doi: 10.1016/j.scib.2020.09.001
[11] Li H, Arculus R J, Ishizuka O, et al. Basalt derived from highly refractory mantle sources during early Izu-Bonin-Mariana arc development [J]. Nature Communications, 2021, 12: 1723. doi: 10.1038/s41467-021-21980-0
[12] Sasaki T, Yamazaki T, Ishizuka O. A revised spreading model of the West Philippine Basin [J]. Earth, Planets and Space, 2014, 66: 83. doi: 10.1186/1880-5981-66-83
[13] 吴时国, 范建柯, 董冬冬. 论菲律宾海板块大地构造分区[J]. 地质科学, 2013, 48(3):677-692 doi: 10.3969/j.issn.0563-5020.2013.03.008 WU Shiguo, FAN Jianke, DONG Dongdong. Discussion on the tectonic division of the Philippine Sea Plate [J]. Chinese Journal of Geology, 2013, 48(3): 677-692. doi: 10.3969/j.issn.0563-5020.2013.03.008
[14] Mrozowski C L, Lewis S D, Hayes D E. Complexities in the tectonic evolution of the West Philippine Basin [J]. Tectonophysics, 1982, 82(1-2): 1-24. doi: 10.1016/0040-1951(82)90085-3
[15] Hilde T W C, Lee C S. Origin and evolution of the west Philippine Basin: A new interpretation [J]. Tectonophysics, 1984, 102(1-4): 85-104. doi: 10.1016/0040-1951(84)90009-X
[16] 张国良, 王帅, 张吉, 等. 西太平洋若干沟-弧-盆体系及板内岩浆成因研究进展[J]. 海洋与湖沼, 2017, 48(6):1220-1234 ZHANG Guoliang, WANG Shuai, ZHANG Ji, et al. Proceedings of subduction system and intra-oceanic volcanism of the western Pacific [J]. Oceanologia et Limnologia Sinica, 2017, 48(6): 1220-1234.
[17] Ozima M, Kaneoka I, Ujiie H. 40Ar-39Ar age of rocks, and the development mode of the Philippine Sea [J]. Nature, 1977, 267(5614): 816-818. doi: 10.1038/267816a0
[18] Sutter J F, Snee L W. K/Ar and 40Ar/39Ar dating of basaltic rocks from Deep Sea Drilling Project Leg 59 [J]. Am. Geophys. Union, 1978, 59: 729-734.
[19] Mrozowski C L, Hayes D E. The evolution of the Parece Vela Basin, Eastern Philippine Sea [J]. Earth and Planetary Science Letters, 1979, 46(1): 49-67. doi: 10.1016/0012-821X(79)90065-7
[20] Mills W. Analysis of conglomerates and associated sedimentary rocks of the Daito Ridge, Deep Sea Drilling Project Site 445[R]. Init. Rep. Deep Sea Drill. Proj. 1980: 643-657.
[21] Hall C E, Gurnis M, Sdrolias M, et al. Catastrophic initiation of subduction following forced convergence across fracture zones [J]. Earth and Planetary Science Letters, 2003, 212(1-2): 15-30. doi: 10.1016/S0012-821X(03)00242-5
[22] Murauchi S, Den N, Asano S, et al. Crustal structure of the Philippine Sea [J]. Journal of Geophysical Research, 1968, 73(10): 3143-3171. doi: 10.1029/JB073i010p03143
[23] Rodnikov A G. Deep structure of the tectonosphere in the West Equatorial Pacific margin [J]. Journal of Southeast Asian Earth Sciences, 1991, 6(3-4): 201-208. doi: 10.1016/0743-9547(91)90067-8
[24] Nishizawa A, Kaneda K, Katagiri Y, et al. Variation in crustal structure along the Kyushu-Palau Ridge at 15-21°N on the Philippine Sea plate based on seismic refraction profiles [J]. Earth, Planets and Space, 2007, 59(6): e17-e20. doi: 10.1186/BF03352711
[25] Uyeda S, Ben-Avraham Z. Origin and development of the Philippine Sea [J]. Nature Physical Science, 1972, 240(104): 176-178. doi: 10.1038/physci240176a0
[26] Karig D E. Origin and development of marginal basins in the western Pacific [J]. Journal of Geophysical Research, 1971, 76(11): 2542-2561. doi: 10.1029/JB076i011p02542
[27] Hickey-Vargas R. Basalt and tonalite from the Amami Plateau, northern west Philippine Basin: New Early Cretaceous ages and geochemical results, and their petrologic and tectonic implications [J]. The Island Arc, 2005, 14(4): 653-665. doi: 10.1111/j.1440-1738.2005.00474.x
[28] Ishizuka O, Tani K, Reagan M K, et al. The timescales of subduction initiation and subsequent evolution of an oceanic island arc [J]. Earth and Planetary Science Letters, 2011, 306(3-4): 229-240. doi: 10.1016/j.jpgl.2011.04.006
[29] Tani K, Ishizuka O, Ueda H, et al. Izu-Bonin arc: intra-oceanic from the beginning? Unraveling the crustal structure of the Mesozoic proto-Philippine Sea Plate[C]//Proceedings of the AGU Fall Meeting 2012. Washington, D. C. : AGU, 2012.
[30] Hall R. The subduction initiation stage of the Wilson cycle [J]. Geological Society, London, Special Publications, 2019, 470(1): 415-437. doi: 10.1144/SP470.3
[31] Tokuyama H. Origin and development of the Philippine Sea[M]//Tokuyama H, Sheheka N, Isezaki N, et al. Geology and Geophysics of the Philippine Sea. Tokyo: Terrra Science Publisher, 1995: 155-163.
[32] Matsuda J, Zashu S, Oziam M. Sr isotope studies of volcanic rocks from island arcs in the western Pacific [J]. Tectonophysics, 1977, 37: 141-151. doi: 10.1016/0040-1951(77)90044-0
[33] Deschamps A, Monié P, Lallemand S, et al. Evidence for Early Cretaceous oceanic crust trapped in the Philippine Sea Plate [J]. Earth and Planetary Science Letters, 2000, 179(3-4): 503-516. doi: 10.1016/S0012-821X(00)00136-9
[34] Qian S P, Zhang X Z, Wu J, et al. First identification of a Cathaysian continental fragment beneath the Gagua Ridge, Philippine Sea, and its tectonic implications [J]. Geology, 2021, 49(11): 1332-1336. doi: 10.1130/G48956.1
[35] Hall R, Nichols G J. Terrane amalgamation in the Philippine Sea margin [J]. Tectonophysics, 1990, 181(1-4): 207-222. doi: 10.1016/0040-1951(90)90017-3
[36] Yumul G P Jr. Westward younging disposition of Philippine ophiolites and its implication for arc evolution [J]. Island Arc, 2007, 16(2): 306-317. doi: 10.1111/j.1440-1738.2007.00573.x
[37] Lewis S D, Hayes D E, Mrozowski C L. The origin of the West Philippine Basin by inter-arc spreading[M]//Balce G R, Zanoria A S. Geology and Tectonics of the Luzon-Marianas Region: Philippine Seatar Committee Special Publication, 1982, 1: 31-51.
[38] Lallemand S, Arcay D. Subduction initiation from the earliest stages to self-sustained subduction: Insights from the analysis of 70 Cenozoic sites [J]. Earth-Science Reviews, 2021, 221: 103779. doi: 10.1016/j.earscirev.2021.103779
[39] Stern R J. Subduction initiation: spontaneous and induced [J]. Earth and Planetary Science Letters, 2004, 226(3-4): 275-292. doi: 10.1016/S0012-821X(04)00498-4
[40] Shemenda A I. Horizontal lithosphere compression and subduction: constraints provided by physical modeling [J]. Journal of Geophysical Research:Solid Earth, 1992, 97(B7): 11097-11116. doi: 10.1029/92JB00177
[41] Deschamps A, Lallemand S. The West Philippine Basin: An Eocene to early Oligocene back arc basin opened between two opposed subduction zones [J]. Journal of Geophysical Research:Solid Earth, 2002, 107(B12): 2322.
[42] Leng W, Gurnis M. Subduction initiation at relic arcs [J]. Geophysical Research Letters, 2015, 42(17): 7014-7021. doi: 10.1002/2015GL064985
[43] Ribeiro J M, Stern R J, Kelley K A, et al. Composition of the slab-derived fluids released beneath the Mariana forearc: evidence for shallow dehydration of the subducting plate [J]. Earth and Planetary Science Letters, 2015, 418: 136-148. doi: 10.1016/j.jpgl.2015.02.018
-
期刊类型引用(2)
1. 宋婉婷, 黄宝琦. 浮游有孔虫壳体钙化的影响因素及壳体重量研究进展. 微体古生物学报(中英文). 2025(02)  百度学术
百度学术
2. 秦秉斌,熊志方,贾奇,郭景腾,李铁刚. 浮游有孔虫Neogloboquadrina dutertrei钙化作用及其海洋酸化指示意义. 第四纪研究. 2025(02): 588-599 . 百度学术
其他类型引用(0)
![图 1 全球热带大洋表层沉积物P. obliquiloculata壳体重量与深海Δ[CO32-]的关系[14]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2022/5/2023071001-1.jpg)
 下载:
下载:
![图 2 全球热带大洋(A)以及单一海区中(B)表层沉积物N. dutertrei壳体重量与深海Δ[CO32-]的关系[14]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2022/5/2023071001-2.jpg)
![图 3 全球热带大洋(A)及单一海区中(B)表层沉积物T. sacculifer壳体重量与深海Δ[CO32-]的关系](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2022/5/2023071001-3.jpg)
![图 4 南大洋南极带表层沉积物N. pachyderma(sin.)壳体重量与深海Δ[CO32-]的关系[17]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2022/5/2023071001-4.jpg)
![图 5 T. sacculifer和N. dutertrei的壳体重量-深海Δ[CO32-]的经验校准公式[28]](/fileHYDZYDSJDZ/journal/article/hydzydsjdz/2022/5/2023071001-5.jpg)